Few of us need an introduction to the Crystal Palace dinosaur sculptures (here shown during construction in 1854), or their artist, Benjamin Waterhouse Hawkins, but not many of us know much about Hawkins or his work beyond this one installation, thus overlooking a major contributor and pioneer in the early history of palaeoart. Image from the Friends of Crystal Palace Dinosaurs blog.
Most of us would agree that palaeoart history is marked by a few key figures that, for one reason or another, define their generation. Knight, Burian, and Paul are perhaps the most considered of these, each producing an iconic portfolio of work that set the bar for other artists and recast how we might view and consume palaeoartworks. But the professional grandfather to all these famous figures is Benjamin Waterhouse Hawkins, a familiar name to anyone interested in palaeoart or vertebrate palaeontology, and yet one that doesn't always receive the accolade and praise it perhaps deserves. Hawkins - whose life spanned 1807-1894 - became the first great palaeoartist at a time when both palaeoart and palaeontology was still finding their feet. We all know of Hawkins' work at Crystal Palace, where he built 33 life-sized restorations of prehistoric mammals, reptiles and amphibians as part of the grand educational and commercial extravaganza, but many of us do not know much about him, his other works, or even how groundbreaking and unique his Crystal Palace sculptures were, demonstrating the sort of good practise and insight that we regard as essential in modern palaeoart.
Image may be NSFW. Clik here to view.
Image may be NSFW. Clik here to view.
Image may be NSFW. Clik here to view.
Hawkins was a skilled anatomist who wrote several books on animal anatomy and form. These images, from his 1860 book A comparative view of the human and animal frame, show his mastery of osteology and anticipate the now-common convention of a dark soft-tissue silhouette behind our skeletal reconstructions.
An appreciation of Hawkins is helped by learning something of his life. Although featuring frequently in accounts of Victorian-age natural history, the details of Hawkins' life had not been pieced together in real detail until his great great, great-granddaughter Valerie Bramwell and historian Robert M. Peck synthesised his personal and professional records into a concise biography (Bramwell and Peck 2008). Hawkins' existence was an eventful one, including both great professional success and tragedy, as well as a complex and somewhat mysterious personal life. The latter is perhaps the most cryptic and unexpected part of the Hawkins story, as he somehow sustained nearly four decades of bigamy (juggling two wives and 10 children, seven of whom survived infancy) without his families discovering the deception. His strange domestic life was found out in the mid-1870s, not long before a string of personal tragedies and a stroke brought Hawkins' life to a sad, inauspicious end. But despite his bizarre private dealings, Hawkins seems to have been a well-liked, generous man known as hardworking, personable and charming, equally confident of his abilities as an artist and anatomist but also reverential and respectful to his peers, especially the academics and intellectuals he frequently worked with.
Hawkins never secured permanent or long-lasting employment but instead moved from commission to commission and project to project. Though he dabbled in a few other careers, his chief profession was creating art of animals - initially living, and eventually extinct. Known best today for his sculpture, Hawkins only began working in 3D during the 1840s, well after he had established himself as an expert painter and illustrator. He was regarded as a high-quality, experienced natural history artist early in his career such that, even before he reached his career peak - the Crystal Palace geological restorations - he was sought-after as an illustrator of zoological specimens. He produced art for many of the biggest names in contemporary palaeontology and biology, including William Buckland, Gideon Mantell, Richard Owen, Thomas Henry Huxley, Charles Darwin and Joseph Leidy (interestingly, Hawkins could have worked with another big name, Edward Cope, but he disliked and refused to work with him - see Desmond 1976). Hawkins' association with Darwin and Huxley is of interest because he became an outspoken anti-evolutionist in later life, a position he expressed directly and clearly in many of the popular public lectures he delivered in his post-Crystal Palace career.
Image may be NSFW. Clik here to view.
Before Hawkins recreated extinct dinosaurs, he was a prolific and sought-after illustrator of the modern kind, along with many other types of animals. This illustration of a White-crested Kalij was produced for John Edward Gray's Illustrations of Indian Zoology (1830-1835). (From Bonhams; incidentally, original Hawkins' lithographs like this sell for hundreds or even thousands of pounds to private collectors, a sad contrast with the under-financed conservation of his Crystal Palace works.)
Hawkins was well-known for his expertise in animal anatomy and form, which he learned primarily from zoological museum specimens as well as drawing captive animals (Bramwell and Peck (2008) recount an urgent trip to Paris in 1849 to sketch a newly born giraffe calf). As would happen later with Charles Knight, this expertise with living animals was instrumental in his later work at reconstructing extinct ones. Hawkins authored several books on animal anatomy and from the 1850s onward spent a large amount of time - and commanded a high fee - for grand public lectures. At his lecturing acme, Hawkins produced large drawings of prehistoric animals on stage (of such size that a ladder was required to reach the top of the board canvas) to bring his restorations direct to his audience. A gleaming reputation among British academics meant he enjoyed a warm reception in the US where he was offered many auspicious platforms and opportunities by well-regarded officials and academic institutions. These included the production of (sadly never realised) Crystal Palace-like prehistoric animals installations in Central Park and the Smithsonian, and production of the first-ever mounted dinosaur skeleton (Hadrosaurus), of which several casts were made (of which only one skull survives).
Hawkins' Central Park Workshop, c. 1869, with an assembled Hadrosaurus mount and the beginnings of a Laelaps skeletal reconstruction alongside - note the mount outline to the right of the Hadrosaurus ;skeleton. This terrific image (reproduced from Bramwell and Peck 2008) is a treasure trove of detail: look out for various mouldings of reptile skin, including a carnivorous species on the left, real animal skeletons in the background and unmounted bone replicas on the floor (are these bits of Laelaps?). There's also a lack of vermin - Hawkins' Crystal Palace workshop was shown with rats and birds scampering over the floor, and described as lying among a muddy swamp. The content of this workshop was destroyed by order of corrupt New York politicians, which devasted Hawkins.
There's a lot more we could say about Hawkins career, but you get the point: Hawkins wasn't a flash-in-the-pan artist who happened to land the Crystal Palace gig, nor was he someone who just knew how to throw clay and concrete around to someone else's design. He was a distinguished and respected intellect with expertise in natural history and anatomy, and brought considerable experience to his palaeoart commissions. Indeed, historians are uncovering increasing evidence that Hawkins was the principle intellect behind many of the Crystal Palace sculptures. We touched on this in my previous discussions of the Crystal Palace palaeoartworks but it's worth repeating: Richard Owen, lauded by many (e.g. Phillips 1854; Owen 1894; Desmond 1979; Beaver 1986; McCarthy and Gilbert 1994) as the brains and overseer of the Crystal Palace models, was actually barely involved and may have even held disdain for this grand project (Secord 2004; Dawson 2016). Numerous pieces of evidence, the best of which are the private correspondence from Hawkins, Crystal Palace officials and others, show that Owen only visited the models once during their construction for the famous 1853 New Years's Eve banquet held in the clay Iguanodon. Most tellingly, Hawkins is on record as stating that Owen "afforded no assistance" while the models were being built (Dawson 2016). Owen later attempted - as was his custom - to take greater credit for his role in the project, but his actual contributions seem restricted to input on some early clay maquettes constructed by Hawkins; a short, incomplete guide book about the sculptures (Owen 1854); some promotional duties; and allowing his name to be used to give the restorations scientific authenticity. We can thus draw two conclusions: 1) having been largely abandoned by his consultant, Hawkins' expertise and anatomical confidence were probably instrumental in seeing the models realised and completed on time; and 2) the sculptures are not - as so often stated - Owenian theory brought to life, but Hawkins' personal take on the works of Owen, Mantell and Cuvier. This probably explains why, as we discussed previously, many details of the models are at odds with Owen's ideas, as well as the nitpicking tone his 1854 sculpture guidebook.
Image may be NSFW. Clik here to view.
Hawkins' 1870s take on the moa, produced for College of New Jersey (now Princeton University). The moa is not a creature we intimately associate with Hawkins, but its research history - where its form was predicted from a small amount of fossil material - underpinned Hawkins career as a palaeoartist. From Wikipedia.
It's in this largely unguided context that we have to view many of the details in Hawkins' Crystal Palace work as the product of a genuine palaeoart master, and especially so given their incorporation of an important but ultimately flawed 19th century way of interpreting extinct animals. Hawkins was a student of the then-in vogue Cuvierian philosophy of anatomical correlation: that is, the idea that whole animals could be reconstructed with some degree of precision from very few anatomical remains. This concept, explored in depth by Gowan Dawson in his 2016 book Show Me the Bone, gained traction after successful demonstrations in the early 1800s by Cuvier and then, most famously, by Owen's seemingly miraculous reconstruction of the moa from a femoral fragment. Anatomical correlation became the principle philosophy guiding the earliest visualisations of all poorly known extinct creatures and became celebrated as a scientific marvel in popular and academic spheres alike. Although some scholars - including Mantell - developed misgivings about anatomical correlation by the time the Crystal Palace project began in August 1852, Hawkins relied on this technique to restore his dinosaurs as well as Dicynodon, Mosasaurus and Labyrinthodon, none of which were known from more than scraps of bone at the time. In lieu of a reliable consultant, such predictive restorations might have been disastrous, had they ever been achieved at all, but Hawkins' anatomical expertise instead saw the creation of genuinely lifelike, plausible-looking restorations. Moreover, as discussed in my previous posts, his restorations of Crystal Palace are often more precedent than they first appear, anticipating not only many genuine aspects of prehistoric animal anatomy but also demonstrating sound, logical palaeoart approaches that we can approve of today.
The buffalo-like shoulder hump of Hawkins'Megalosaurus has a slightly complex backstory. Sometimes logically assumed to reference the tall-spined vertebrae of Altispinax dunkeri (a species once part of the Megalosaurus taxonomic complex), it was actually a Hawkinsian speculation that large-skulled dinosaurs needed vast neck and shoulders musculature to support their heads. Initial interpretations of Altispinax seemingly proved Hawkins right, but, of course, we've since realised that the Altispinax vertebrae belong much further down the body, possibly representing a Concavenator-like sail.
Using the dinosaur sculptures as an example of this, it's evident that Hawkins was anticipating the sort of anatomy that giant, somewhat mammal-like giant reptiles might have had to support their vast frames, and was moving well beyond the brief lifestyle and functional commentary provided by Owen and Mantell. He predicted, for instance, that the large heads of dinosaurs would need additional support from their shoulder regions, leading to the portrayal of both Megalosaurus and Iguanodon with shoulder humps of varying size (this is especially obvious in the Megalosaurus, but a shoulder prominence is also discernable in the reclined Iguanodon, while the shoulders of the standing animal bulge into a voluminous, rhino-like neck). This was an entirely sensible prediction that mirrors how we approach reconstructing fossil animals today: when we see robust and large osteological features, we assume they have correspondingly developed soft tissues for support and motion. In this respect Hawkins was ahead of some later artists who essentially ignored aspects of functional morphology and biomechanics in their work, leading to emaciated, peculiarly proportioned animals, such as the emaciated Stout and Kish dinosaur reconstructions from the 1970s and 1980s. Moreover, Hawkins was proved correct in assuming dinosaurs with large heads needed additional neck support, once (as it would turn out, erroneously) during his lifetime when Owen referred the tall-spined dorsal vertebrae of Altispinax dunkeri to the shoulder of Megalosaurus, specifically commenting on their use to support the head (Owen 1856) and, later, by the genuine shoulder and skull anatomy of ceratopsids and certain theropods, which bear augmented anatomy related to skull support.
Hawkins also predicted that dinosaur torsos may differ markedly depending on diet and habits, moreso than they do in conventional reptiles. The herbivorous Iguanodon has a vast belly that spreads in the reclined sculpture, simulating the weight of a large animal resting on a voluminous gut. In contrast, the Megalosaurus has a taut, narrow torso without a significant gut region, more in line with that of a carnivore. At this point in history virtually nothing concrete was known about dinosaur toro dimensions, so these reconstruction choices - also borne out by later discoveries - were sensible predictions of the functional properties of dinosaur guts. A third example concerns his dinosaur musculature, which was modelled after a mammalian, rather than lizard-like fashion. Though mammals are not the best reference for dinosaur myology, Hawkins was right in assuming that large, powerful animals needed large, powerful limbs, and realised that mammals were a more appropriate model than modern reptiles. In this regard, Hawkins was more insightful than some of his successors, who would restore dinosaurs with skinny, lizard-like limbs ill-suited to their (by then well-known and obvious) limb girdles (see artwork by Cope, Knight, Smit etc.). He also went so far as to add features in his art that we associate with especially thorough extinct animal reconstructions today, such as skin webs linking limbs and body, tissue deformation, pose-based muscle bulges and so on.
It's been said before, but it's worth saying again: the reclining Crystal Palace Iguanodon is an amazing work of animal reconstruction. This 30-tonne concrete and brick model captures minutiae of superficial musculature and other nuanced features of anatomy. The world had not seen anything like this in 1854, and even today it knocks the stuffing out of the detailing seen on many dinosaur sculptures.
These points demonstrate how excellent Hawkins was at solving anatomical 'problems' thrown up by fossil remains with an appropriate and sensible corresponding morphology. They belie an experience and instinct with anatomy and biological functionality that remains essential to high-quality palaeoart today, and this was undoubtedly a major factor in how these largely speculative creatures look so realistic. Hawkins' predictions were not always correct of course, but his decisions were logical given the material available to him - the best we can aim for in any palaeoartwork, regardless of its vintage. Needless to say, plenty of people were amazed by his work, and both its popularity and seeming scientific authenticity saw it referenced and replicated by artists for decades to come.
Because Hawkins is mainly known for the Crystal Palace models few people realise that he enjoyed a significant post-Crystal Palace career revolving around reconstructing extinct life. His skills were sought out by various bodies for use in museums, posters and other artworks. During lengthy visits to the United States, he almost re-realised the grandeur of the Crystal Palace models (first in Central Park, later the Smithsonian), but circumstances were never on his side and these projects never materialised or - in the case of the Central Park Paleozoic museum - were sabotaged. Hawkins continued to produce palaeoart until at least the 1870s where he completed what could be considered as his second most substantial set of palaeoartworks: 17 paintings of different geological periods for the College of New Jersey (now Princeton University). These artworks capture the evolution of Hawkins' personal ideas on prehistoric life and are also worth tracking down as exemplary takes on prehistory from the late 19th century. Among the most interesting is the 1877 painting Jurassic Life of Europe where Hawkins revisited his Crystal Palace subjects for (so far as I know) the final time. Hawkins got a lot of professional mileage from the Crystal Palace project and many of his subsequent artworks referenced his sculptures in detail, down to their posing, but by 1877 he must have realised that many of his reconstruction choices were no longer tenable. Both the now-hornless Iguanodon and Megalosaurus bear visibly short forelimbs, long, bird-like hindlimbs, and relatively slender necks with somewhat smaller, less bulky heads. Although still quadrupedal, these restorations indicate the influence of new dinosaur discoveries on Hawkins' classic dinosaur interpretations. This 1877 work might be criticised for his animals not having attained the status of true bipeds, as Hawkins realised was appropriate for both Hadrosaurus and "Laelaps", but consider that Megalosaurus was still considered to have powerful, bulky shoulders at this time (a holdover from Owen's interpretations) and that, while Iguanodon was considered by Mantell (1848) to have relatively gracile limbs capable of non-supportive functions (e.g. grasping vegetation), an explicit case for Iguanodon bipedality had not yet been made - the famous Bernissart Iguanodon skeletons would not be found until a year after Hawkins completed his painting, in 1878. Jurassic Life of Europe is surely among the last palaeoartworks where overtly Mantellian and Owenian interpretations of dinosaurs could be said to have some validity.
Image may be NSFW. Clik here to view.
Hawkins' 1877 painting Jurassic Life of Europe, produced for the College of New Jersey (from Princeton University Art Museum).
Jurassic Life of Europe and its sister paintings demonstrate Hawkins' adaptability and humility as a palaeoartist. Much of Hawkins fame came from the Crystal Palace Company promoting their dinosaurs as authentic, wholly accurate creations, so their adaption to fit more modern ideals was an admission that his most revered work was not without some significant errors, and that the predictive principle at the heart of the geological court was not infallible. Concerns that the Crystal Palace sculptures were flawed were not new at this point (Dawson 2016) but, by the late 19th century, they were subject to increasing vicious comments from detractors. Othniel Marsh, speaking in 1895, remarked
The dinosaurs seem... to have suffered much from both their enemies and their friends. Many of them were destroyed and dismembered long ago by their natural enemies, but, more recently, their friends have done them a further injustice by putting together their scattered remains, and restoring them to supposed lifelike forms... So far as I can judge, there is nothing like unto them in the heavens, or on the earth, or in the waters under the earth. We now know from good evidence that both Megalosaurus and Iguanodon were bipedal, and to represent them as creeping, expect in their extreme youth, would be almost as incongruous as to do this by the genus Homo.
Othniel C. Marsh, 1895 (quoted in Desmond 1976)
Marsh and other late 19th century critics were, of course, not being entirely fair to Hawkins by comparing his work to their contemporary knowledge of prehistory, and not judging it by the standards of decades prior. Perhaps, for those living through the rapid paleontological advancements of the late 19th century, Hawkins' attempts to reconstruct fossil animals from fragmentary remains, and the genuine belief that they were credible takes on their true form, seemed premature, arrogant and foolish. There may be some truth to this: the 19th century acceptance of anatomical correlation is surely a major case of Dunning-Kruger effect, where scholars had yet to be humbled by the bewildering anatomical diversity present in Deep Time and believed their mastery of modern natural history equipped them to make bold, confident predictions about the past. But it's surely also the case that, while the science of his time was flawed, Hawkins' work was as true to that science as could realistically be expected. Like the kids say, don't hate the player.
Image may be NSFW. Clik here to view.
The final extinction of Hawkinsian dinosaurs: the frontispiece to Camille Flammarion's Le monde avant la création de l'homme& (1886). In this (possibly) last use of Hawkins' restorations in a non-historic, non-ironic context, a Crystal Palace Iguanodon (right) meets an early interpretation of Stegosaurus (left) - a representation of New World dinosaurs coming into focus. By this time, new European fossils had already made Hawkins' reconstructions long in the tooth, and American specimens were showing how wide of the mark his predictions were.
Hawkins' death in 1894 occurred without note, despite his significant contributions to science, public outreach, and education. Thankfully, his legacy has not gone neglected and increasing research into his life and work has seen his stock as an artist and intellect grow considerably, at least among academics. But it remains the case that showcasing Hawkins' work to even palaeoart fans sees it quickly dismissed, as by Marsh, as inaccurate, and thus now worthless, takes on prehistory. It's surely the case, however, that drawing attention to the inaccuracies of Hawkins' palaeoart is the most superficial and least interesting observation one can make of it, and defies the obvious fact that palaeoart of any kind must be evaluated in an appropriate historic context. Of course Hawkins' work is inaccurate, and pointing it out does not make one look smart or insightful. Rather, look closely and we see the work of someone who, with no prior experience in the prehistoric animal restoration, took the early palaeaort into a different league of quality and popularity; pioneered principles of soft-tissue restoration and attention to detail that we consider essential today; and demonstrated that even the most unusual extinct animals, with sufficient knowledge of living species, could be restored to plausible, life-like forms. The outdated science underlying Hawkins' work may not it the most accessible of vintage palaeoart, but it's quality, the knowledge that informed it, and historic significance leaves little doubt that Hawkins was a true master of the genre, and worthy of deeper study and appreciation by palaeoart enthusiasts.
Postscript
Image may be NSFW. Clik here to view.
In June 2020 the face of the Crystal Palace Megalosaurus, surely one of the most iconic works of Victorian palaeoart on the planet, was significantly damaged. The survival of these works, and Hawkins' legacy, is a continued fight against the elements and the thoughtless people who routinely clamber over them. Image from the Friends of Crystal Palace Dinosaurs website.
Having just written about Hawkins at length, it would be remiss not to mention the terrible damage that recently occurred to the iconic Crystal Palace Megalosaurus, where the front of the face was broken off through suspected vandalism. The Friends of Crystal Palace Dinosaurs charity is now working to repair the damage with Historic England, Bromley Council and specialist conservators, as well as to address issues with the security of the island that houses most of Hawkins' sculptures. Note that this incident has nothing to do with the new bridge to the island, which has not yet been installed and, in any case, will be a rotating structure inaccessible to the public most of the time. Instead, the chief security issue is the low water level around the island, making it easy to access. As you can imagine, fixing this damage and ensuring the security of the Crystal Palace models is an expensive undertaking, so please support the charity if you can. If you really think Hawkins' work is worth preserving long-term, you can set up a recurring monthly donation - a small amount each month can quickly build into a substantial contribution, and gives the FCPD more reliable income over time.
References
Beaver, P. (1986). The crystal palace: A portrait of Victorian enterprise. Phillimore & Company.
Bramwell, V., & Peck, R. M. (2008). All in the bones: a biography of Benjamin Waterhouse Hawkins. Academy of Natural Sciences.
Dawson, G. (2016). Show me the bone: Reconstructing prehistoric monsters in nineteenth-century britain and America. University of Chicago Press.
Desmond, A. J. (1976). The hot-blooded dinosaurs: a revolution in palaeontology. Dial Press.
Mantell, G. A. (1848). XIII. On the structure of the jaws and teeth of the Iguanodon. Philosophical Transactions of the Royal Society of London, (138), 183-202.
McCarthy, S., & Gilbert, M. (1994). The Crystal Palace dinosaurs: The story of the world's first prehistoric sculptures. Crystal Palace Foundation.
Owen, R. (1854). Geology and inhabitants of the ancient world (Vol. 8). Crystal palace library.
Owen, R. (1856). The fossil Reptilia of the Wealden Formations. Part III, Megalosaurus bucklandi. Palaeontographical Society. Monographs, 9, 1-26.
Owen, R. (1894). The Life of Richard Owen by His Grandson the Rev. Richard Owen,... with the Scientific Portions: An Essay on Owen's Position in Anatomical Science by the Right. John Murray.
Phillips, S. (1854). Guide to the Crystal Palace and park. Crystal Palace Library.
Secord, J. A. (2004). Monsters at the crystal palace. In: de Chadarevian, S, & Hopwood, N. (eds). Models: the third dimension of science, Stanford University Press. 138-69.
The American mastodon Mammut americanum is one of the most iconic members of the North American megafauna. A frequent subject of museum displays, books and technical research for over two centuries, we can all immediately conjure mental images of this long-bodied, relatively short-legged elephant-like animal. Nearly all of us will imagine mastodons the same way: that is, covered with a thick layer of brownish hair in a fashion reminiscent of its even more iconic cousin, the woolly mammoth. This is simply how we've come to understand mastodon life appearance through centuries of artistic and literary reinforcement (examples in familiar books include Špinar and Burian 1972; Benton 2015; Prothero 2017). Many texts don't even bother citing academic sources evidencing the claim: the hairy mastodons concept has been repeated often enough and long enough to be established fact. Zebras have stripes, lions have manes, and mastodons had a thick, brown covering of hair.
Zdenek Burian's 1964(?) take on the American mastodon: an entirely typical restoration of this species that would pass as a credible restoration at any point during the last two centuries.
It might come as a shock, therefore, to learn that the foundation of evidence behind our shaggy mastodons was actually entirely baseless for almost two centuries, and that this widely accepted concept has only gained a small amount of support within our lifetimes. This isn't a new revelation, either. American anthropologist and author Loren C. Eisley, a key figure in unravelling the strange history of mastodon soft-tissues, held no punches when describing mastodon hair as "items of palaeontological folklore" in 1945 (p. 108). Eisley's interest in mastodons was driven by ideas of their survival into recent centuries, where so-called mastodon soft-tissues discovered in the 19th century were interpreted as evidence of mastodons dying out just hundreds of years ago. Eisley published rebuttals to this concept several times during the 1940s and, in his 1946 Science article Men, Mastodons, and Myth, he specifically delved into the peculiar history of mastodon soft-tissue discoveries. Through historical detective work, Eisley uncovered a series of erroneous interpretations, failed corrections, and even deliberate subterfuge from the early days of American fossil exploration. A brief summary of his findings are provided below, but be sure to check out Eisley's account yourself for the full picture.
Lies, damned lies, and mastodon hair
The story of mastodon soft-tissue begins in 1800 Newburgh, New York, where a mastodon tooth and associated sample of coarse, 'dun brown' hair was extracted from a bog on the farm of a Mr. A. Colden. This specimen, said to have been so rotten that it decayed to dust within days, was a significant find. It was not only the first alleged mastodon hair, but also the first indication that some Pleistocene giants might have been covered in fur. It's among the more credible accounts of mastodon hair from the 19th century and, for many, was the best evidence of mastodon skin even as new finds were made later in the same century.
Charles Knight's awesome 1897 Mammut americanum painting. So far as I can tell, Knight's work is among the earliest restorations of this species and establishes the shaggy coat we'd come to associate with it. This is one of my favourite Knight paintings: just look at that landscape.
Shortly after the Colden Farm discovery, reports of mastodon soft-tissue then came thick and fast. In 1805 a mastodon was reported by Shawnee native Americans as having a long nose and mouth - interpreted, naturally, as a fossilised trunk. A report of a fossil mastodon stomach with gut content was reported at around the same time, as were more specimens with small amounts of hairy skin. Especially large sheets of skin were reported in 1839. These were reportedly so well preserved that they included arteries and sinew, but were too fragile to collect in one piece - only small fragments could be extracted. Collectively, such remains formed a significant dataset regarding mastodon soft-tissues, from internal organs to external features. They are the seed from which the concept of hairy, brown mastodons grew, and by the end of the century hairy mastodons were described in textbooks (e.g. Hutchinson 1893 - note that Hutchinson's book contains a rare Joseph Smit illustration of a nearly hairless mastodon) and featured in influential Charles Knight artworks (above),
But if this evidence is so great and extensive, why are none of these specimens better known? Are they not on display in museums, or at least illustrated in a book or paper? It turns out that there's a good reason you've never seen them: they're all completely bogus. Not a single example of mastodon soft-tissue reported in the 19th century was accessioned to a museum, virtually none were examined by individuals with paleontological expertise, and no detailed reports were ever made. Some clearly never existed and those that did were almost certainly erroneous takes of otherwise unremarkable objects.
Eisley's investigation assumes that some of these misinterpretations were honest mistakes by inexperienced parties, or perhaps over-interpretations of field reports (e.g. the 'long nose' specimen account makes no actual mention of soft-tissues, and could pertain to osteological observations alone). He attributes these errors to the early 1800s being a time of great excitement about the then newly-discovered Siberian frozen mammoths, and anticipation that fossil American proboscideans would return remains of similar quality. This might explain why some cases, such as with the alleged mastodon stomach specimen, are just outright strange. Gut tissues are among the first organs to decay when animals die and it would be very odd for a stomach to survive, alone, after the rest of the animal had rotted away. This discovery sounds a lot like the sort of over-zealous interpretation that might be made by someone naive about taphonomy and, indeed, this specimen was quickly subjected to rebuttals and corrections from more experienced scholars. ;
But other accounts were likely dishonest from the start - tall-tales to excite interest in fossil specimens set for display in private museums and touring shows. The early 19th century was a time before public museums existed in the USA, so fossil remains were exhibited to the public through private enterprise: hype and publicity-seeking were important to making such efforts financially viable. It's in this context that we have to view the 1839 Missouri mastodon reported to have extensive amounts of preserved skin, arteries and sinew, which was reported by non-other than Albert Koch: the same museum proprietor responsible for reconstructing Basilosaurus as a sea serpent and a Missouri mastodon as the 'Missourium' - a monstrous assemblage of mastodon and wooden blocks toured for public display in the mid-1800s. Koch's exaggerating, fraudulent approach to the conveyance of paleontological data robs all credibility from his accounts, and no-one can take his unverified claim of giant skin samples seriously. The fact that he never followed up on these seemingly remarkable soft-tissue remains is further evidence that they never existed.
Albert Koch's "Missourium" - a composite and distorted mastodon skeleton augmented with bits of wood to make the vertebral column longer. The Missourium specimen was collected around 1840, and I wonder if Koch's accounts of giant skin samples were associated with its discovery.
But what of the Colden Farm find which - while still entirely anecdotal - at least mentioned the unstable nature of its mastodon hair, and thus accounted for its absence in current collections? This 1800 discovery gains additional credibility in pre-dating our knowledge of frozen mammoths in Russia, and must therefore have been an unbiased, honest interpretation of alleged mastodon material. But, again, particulars of this find are peculiar: isn't it strange that an entire mastodon disappeared to leave only a solitary tooth and a patch of hair behind? This is taphonomically very odd, but was not an isolated incident: other fragmentary bones with patches of hair were also recovered from swamps in the same area. The American geologist James Hall provided an explanation of such sites which is far more consistent with our understanding of animal decay patterns. As part of a wider survey of the geology of New York state, Hall found the swamps yielding these fragmentary mastodon remains were filled with a hair-like algae known as conferva. It was a good match, size and morphology wise, for the alleged mastodon hair, and when desiccated, it turned - you guessed it 'dun brown'. When describing a mastodon site in 1843, he wrote:
In a small muck swamp in Stafford, Genesee County, a small molar tooth was found several years since. Its situation was beneath the muck, and upon a deposit of clay and sand. A large quantity of hair-like confervae, of a dun brown color, occurs in this locality; and so much does it resemble hair, that a close examination is required to satisfy one's self of its true nature.
Hall 1843 (from Eisley 1946, p. 522)
Hall was one of the few truly experienced and qualified individuals to write about mastodon soft-tissues in the 19th century, so his assessment is of real interest to this story. This is not to discount the insight of the pastors, farmers and businessmen behind other accounts, but Hall's explanation certainly sits better with our modern understanding of taphonomy, as well as the fact that those New York swamps have - even today - yet to yield a single scrap of incontrovertible mastodon hair. For Eisley, if there's any truth to these early takes on mastodon hair, Hall nailed it: the New York mastodon hair samples were simply misidentified dry algae.
'Conferva' is not a widely used term nowadays, but once pertained to a great number of filamentous green algae species. It's easy to see how examples like the above might be mistaken for mastodon hair by naive parties. Image by Anne Dixon, from 1843-45, borrowed from Getty Museum.
Without any specimens to examine, we cannot be certain today whether Hall and Eisley were correct, but their work clearly shows that 19th century claims of mastodon hair are suspicious. This is the line taken by at least some modern authors writing about mastodon hair (e.g. Hallin 1989; Haynes 1991; Larramendi 2015) but, as we know from history, most people ignored both Hall in the 19th century and Eisley in the 20th to perpetuate the discredited concept of hairy mastodons. How did such questionable data become the established, unquestioned truth about mastodon life appearance? Eisley (1946) attributed this to the illusory truth effect, where the repetition of a claim by perceived authorities makes it seem factual and truthful, regardless of the underlying evidence. In this case, enough scientists, museums, books and other media have towed the hairy mastodon line to transform folklore into 'fact', seemingly without anyone wondering where the real evidence of mastodon hair was. As Eisley put it:
In the midst of this constant repetition of what, through the sheer prestige of age, has come to be accepted as undeniable fact, it has never been pointed out that American institutions of science do not possess the tangible evidence which alone could justify such wholehearted faith in the exact appearance of this long-vanished beast.
Eisley 1946, p. 517.
I struggle to think of a case where interpretations based on a comparably feeble palaeontological dataset have been rehashed so uncritically for so long, so regularly and so publically. There is ordinarily some pushback against wholly baseless ideas of extinct animal life appearance, even if only among specialists, but I can only find a handful of articles promoting non-hairy takes on mastodon life appearance from the last century. In light of dedicated efforts by the likes of Eisley to set the record straight - published in Science, no less - it's really quite baffling that we've unwaveringly promoted hairy, brown mastodon for so long.
Finally: a real specimen
The situation around mastodon soft-tissues has changed somewhat today. In the 1980s, Pleistocene mammal expert Kurt Hallin published two abstracts and one popular article about the first genuine chunks of mastodon skin, both of which were covered in hair (Hallin and Gabriel 1981; Hallin 1983, 1989). But if you're hoping that this finally gives us real insight into mastodon life appearance, you're out of luck. To my knowledge, this specimen has never been described or illustrated beyond these short works, and a scanning electron microscope image of a single hair is all that's been figured of it (below). This short paragraph by Haynes (1991) provides one of the more detailed overviews I could find:
Specimens of what appears to be carbonized skin holding together bundled and fine hairs interspersed with hollow, coarser hairs may be the only preserved Mammut soft tissue currently known. These specimens were recovered by Krut Hallin in association with cranial fragments found near Milwaukee, Wisconsin (K. Hallin 1989 personal communication). The preserved guard hairs are hollow, a common enough trait in mammals, including woolly mammoths and African elephants. The underfur appears similar to that of semi-aquatic mammals such as the otter and beaver (Hallin 1983, 1989; Hallin and Gabriel 1981), in that it is very fine and wavy, and grows in dense bundles.
Haynes 1991, p. 34.
30 years on, these brief reports remain our only direct evidence of mastodon hair (Haynes 1991; Larramendi 2015), leaving the life appearance of the American mastodon not significantly advanced from Eisley's dismantling of 19th century discoveries from 75 years ago. While the Wisconsin hair specimen represents a potential step forward, it remains in a scientific grey area for having never being described or illustrated in detail, and Hallin's interpretation has never been subjected to peer review. Its significance to mastodon palaeobiology and life appearance thus remains an open question, as does the nature of mastodon skin in general. After all, the Wisconsin specimen only represents skin from the cranium, and we really need skin from different regions of extinct animal bodies to be confident about their full appearance. ;With such little data to hand, the whole body integument of mastodons remains mysterious (Haynes 1991; Larramnedi 2015).
Hallin's (1989) SEM shot of mastodon hair from Wisconsin - that's it on the left. To my knowledge, this is the only published image of mastodon hair.
Could mastodon still be hairy, though?
But this is not to say that we should just give up on restoring mastodon life appearance, however. These are relatively well-understood fossil animals, and we can surely say something about their skin from details of their anatomy, ecology and palaeobiogeography. If we are to play this predictive game, perhaps the most important first step is to realise that woolly mammoths - which are clearly historic inspiration for mastodon art - are not great analogues for American mastodon. The familiarity of woolly mammoths makes it easy to forget they were genuinely strange, specialist species adapted to extreme cryoarid Pleistocene habitats (Boeskorova et al. 2016). They relied on thick, three-tiered hairy coverings, generous adipose layers, shoulder fat humps, small ears, shortened tails, and trunk tip 'hoods' to protect themselves from extremely cold winters and periodic food shortages (Boeskorova et al. 2016). American mastodon, by contrast, were not Ice Age specialists. They actually evolved during the warmer Pliocene conditions pre-dating the Pleistocene glaciations and, even during the Ice Age, avoided the extreme climates endured by woolly mammoths. The distinct habitat preferences of mastodon and woolly mammoths are demonstrated by their remains being very rarely found in the same fossil horizons (Graham 2001; Hodgson et al. 2008). Woolly mammoths were denizens of steppe, tundra and forest habitats in northern regions (specifically, Alaska, the upper midwestern and northeastern United States, and the northern Atlantic coastal plain), while mastodon preferred wetter woodlands in more southerly locations (the eastern US (extending as far south as Florida), southeastern Canada, and parts of Mexico) (Haynes 1991; Graham 2001; Newsom and Mihlbachler 2006). Mastodon thus enjoyed a range of habitats and climates: northernmost populations inhabited boreal forests, living alongside moose and beavers, while those of Florida and Mexico inhabited relatively balmy swamps and woodlands, sharing their environment with reptiles and amphibians (Hine et al. 2017). Without soft-tissue mastodon fossils we can't truly assess their cold adaptations, but Larramendi (2015) noted that Mammut tails are long for proboscidians. This contrasts with the especially short tails of woolly mammoths and might have implications for mastodon thermal energetics - in other words, they weren't feeling the cold enough to shorten their tails (Larramendi 2015). These details of ecology, biogeography and anatomy demonstrate how different mastodon and woolly mammoths were, and caution against our porting of mammoth skin to their stockier cousins.
Woolly mammoths are very familiar to us, but we should not forget that they were not 'normal' giant animals. They were specialists adapted to life in extremely cold, dry habitats and, like most specialists, their lifestyle required a glut of anatomical weirdness. We should not readily assume that they are suitable model species for other giants. This 2018 image shows M. primigenius meeting a family of Neanderthals.
Having taken mammoths out of the picture, we can focus on a key question about mastodon: would they have benefitted from a full-body covering of hair? To answer this, we have to consider their giant size, and what that means for their thermal energetics. I'm increasingly of the opinion that that, when considering the life appearance of gigantic, multi-tonne extinct animals, we should be justifying the presence of thick layers of insulation, not the removal of it, and I feel that approach has merit here. American mastodon averaged body masses of 8 tonnes, making them larger than not only their distant woolly cousins (Larramendi 2015) but also considerably heavier than an average living elephant of any species. As discussed at length in previousposts, even at these smaller masses, elephants are simply so big that they really don't feel cold that much, to the extent that some African populations endure months of sub-freezing nights despite their lack of hair (Haynes 1991). Indeed, we know that elephants likely rely on these cooler periods to survive, as their size and low surface area-volume ratio present them with numerous challenges related to heat loss during the day. Among other issues, they cannot shed heat as quickly as it is generated during exercise, so their bodies regularly warm to dangerous hyperthermic temperatures. With little direct control over their body temperatures - they cannot even sweat or pant - they rely on features of their environment - bodies of water for drinking, trunk spraying, wallowing, cool nighttime temperatures etc. - to remain cool (Wright and Luck 1984; Weissenböck et al. 2012; Rowe et al. 2013).
Given that mastodon represent an even stockier, heavier and shorter-legged variant on the elephant body plan, they surely faced similar, or even more pronounced, challenges on this front. Cooler Pleistocene temperatures would have alleviated these concerns somewhat, but were unlikely to solve them outright. Elephants generate their own climate envelope around their bodies when exercising so that, even away from their very hot natural ranges, they overheat when exercising for long periods (Rowe et al. 2013). The insulative effects of fibrous integuments compound considerably with body size - a layer of fuzz over a one-tonne animal has a significantly greater insulative effect than the same layer on a 1 kg animal (Fariña 2002; Porter and Kearney 2009). Thus, even if smaller mammals living alongside mastodon required fur coats, mastodon themselves may not have needed them. Based on the thermal tolerances of living animals, we can predict that a blanket of fur all over a mastodon would have a significant impact on their thermal physiology, likely pushing their thermal neutrality (crudely explained, their ambient temperature 'comfort zone') many degrees below zero. Consider that Fariña (2002) calculated thermal neutrality for a naked 4 tonne Megatherium as -17°C*: I not sure what the thermal neutrality of a mastodon is, but their additional 4 tonnes of mass would surely push their thermal neutral figure even lower, maybe much lower. These values are only indicative because they assume dry, windless conditions, and both wind and rain make animals more vulnerable to cold, but they give a sense of how cold-resistant multi-tonne animals are. I'm guessing that Floridan mastodons felt pretty hot a lot of the time.
*Before you ask, this calculation also assumed a placental-typical metabolic rate, so the comparison to mastodon is apt.
Another question we might ask is how prone animals inhabiting generally cold habitats, such as boreal forests, are to heat stress. Does the cold in these settings negate any risk of overheating? Weighing up to 750 kg, moose and elk - members of Alces - are some of the largest modern animals to inhabit boreal forest, and their thermal energetics are well studied. They thus provide useful insight into how large animals cope with cold climates. Winters in boreal forests can get seriously cold - an average day might be -20°C - but moose are actually still prone to heat-stress during these months (Dussault et al. 2004; van Beest and Milner 2013). These aren't just occasional occurrences, either: moose have to manage their heat-stress continually via habitat choice and behavioural modification. If they don't, the associated energy drain to cool their bodies dramatically impacts their mass and health over the winter months (van Beest and Milner 2013). If these cervids find their whole-body fur coats stifling in boreal forest winters, how would mastodon - creatures ten times their size or more - cope with similarly hairy skin? I appreciate that this comparison is very crude - deer are not proboscidians, and modern boreal forests might be climatically different to those of the Pleistocene - but it demonstrates that we should not automatically equate cold climates with cold-stressed animals, and that we should think about the physiological implications of insulating fossil giants, even in frosty settings.
An American mastodon in the cypress swamps of Pleistocene Florida, sporting a hairy face and shoulders, but a largely hairless body. This looks odd compared to our standard fully-hairy restoration, but is consistent with our cranial mastodon hair sample and the thermal physiology of very large proboscideans. Happily, there's enough hair left for an obvious Simpsons gag. Mmm... mastodon fresh.
Putting all this together, there are clear grounds to question whether mastodon needed full hair coats. Consider the evidence: we know that elephant-like mammals already struggle more than other species with heat loss; that an 8 tonne animal is going to have very low thermal neutrality even without a fur coat; that mastodons were not universally denizens of especially cold settings; and that at least some parts of their anatomy - their tails - show opposing conditions to hairy, cold-adapted relatives. Based on this, I'm wondering if parts of the mastodon body - maybe even large portions of it - had reduced or absent hair to negate heat stress (above). This may have been especially important for mastodon living in warmer southern regions or areas with hot summers. Perhaps seasonal or geographic variation in hair growth and distribution was also employed, so that mastodon living in the warm south were almost as hairless as living elephants, and those exposed to cold winters were a little hairier? Of course, we cannot ignore the data from the Wisconsin hair specimen, which shows at least some mastodon faces were hairy, but we should be cautious about extrapolating this to a full-body covering. Proboscidian faces are actually one of the few parts of their bodies which are vulnerable to the cold, as their ears and trunks can develop frostbite (Haynes 1991). The face is thus one of the regions we might expect hair to be present in mastodon enduring cold winters, and it may not reflect the thermal demands of the rest of the body. Clearly, what's needed here is a dedicated study factoring mastodon mass, body surface area, metabolism, and heat loss against a nuanced consideration of Pleistocene climates: there are lots of equivalent studies on other fossil animals, and it would be great to get some real data and investigation on this. Until then, I find Asier Larramendi's (2015) comment on this matter an excellent summary: "...the relatively long tail... and the massive body of M. americanum suggest that the prevalent ideas that these animals were covered with a thick coat of fur are probably exaggerated".
From folklore to parable?
Of course, the main point of this article isn't to explore mastodon appearance in detail, but to stress that the nature of their skin is nowhere near as well-evidenced or understood as we've historically implied. I feel that those of us involved in education, research and palaeoart have a job on our hands to revise our misplaced confidence about mastodon life appearance, and to become more open-minded and explorative about what these iconic animals may have looked like. We should also take a moment to think about how this matter differs from our conventional issues with depictions of fossil animals. While it's not uncommon to bemoan how prehistoric animals are discussed and communicated to the public, we mostly do so because of pop culture influences - a film, TV show or book that perpetuates old tropes or fabricates something ridiculous about a prehistoric species. But there's no sensationalised media behind the perpetuation of hairy mastodons: these misleading depictions have come directly from decades of academic texts, officious museum displays and professionally produced palaeoartworks repeating the same unverified facts over and over, all the while ignoring direct rebuttals of the sketchy data behind this depiction. This one's on us, in other words. Perhaps it's time to turn this the "palaeontological folklore" of mastodon hair into a "palaeontological parable": a story to remind us to check and verify even the most basic and well-known information about our subjects from time to time, to ensure we're actually communicating science, and not rehashing mastodon-sized tall tales.
Enjoy monthly insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Benton, M. J. (2014). Vertebrate palaeontology. John Wiley & Sons.
Dussault, C., Ouellet, J. P., Courtois, R., Huot, J., Breton, L., & Larochelle, J. (2004). Behavioural responses of moose to thermal conditions in the boreal forest. Ecoscience, 11(3), 321-328.
Eiseley, L. C. (1946). Men, mastodons, and myth. The Scientific Monthly, 62(6), 517-524.
Eiseley, L. C. (1945). The mastodon and early man in America. Science, 102(2640), 108-110.
Fariña, R. A. (2002). Megatherium, the hairless: appearance of the great Quaternary sloths (Mammalia; Xenarthra). Ameghiniana, 39(2), 241-244.
Graham, R. W. (2001, October). Late Quaternary biogeography and extinction of proboscideans in North America. In The World of Elephants: Proceedings of the 1st International Congress, Rome, Italy (pp. 16-20).
Hallin, K. F. (1983). Hair of the American mastodon indicates an adaptation to a semiaquatic habitat. In American Zoologist (Vol. 23, No. 4, pp. 949-949).
Hallin, K.F. (1989). Wisconsin's Ice Age tuskers: Ice Age elephants and mastodonts. Wisconsin Academy Review, 35, 6-10.
Hallin, K.F. & Gabriel, D. (1981). The first specimen of mastodon hair. Geological Society of America 34th Annual Meeting of the Rocky Mountain Section, Abstracts with Program. 13(4), 199.
Haynes, G. (1991). Mammoths, mastodonts, and elephants: biology, behavior and the fossil record. Cambridge University Press.
Hine, A. C., Martin, E. E., Jaeger, J. M., & Brenner, M. (2017). Paleoclimate of Florida. Florida's Climate: Changes, Variations, & Impacts.
Hutchinson, H. N., & Woodward, H. (1893). Extinct monsters. Chapman & Hall.
Hodgson, J. A., Allmon, W. D., Nester, P. L., Sherpa, J. M., & Chiment, J. J. (2008). Comparative osteology of late Pleistocene mammoth and mastodon remains from the Watkins Glen site, Chemung County, New York. Mastodon Paleobiology, Taphonomy, and Paleoenvironment in the Late Pleistocene of New York State: Studies on the Hyde Park, Chemung, and North Java Sites. Palaeontographica Americana, 61, 301-367.
Larramendi, A. (2015). Shoulder height, body mass, and shape of proboscideans. Acta Palaeontologica Polonica, 61(3), 537-574.
Porter, W. P., & Kearney, M. (2009). Size, shape, and the thermal niche of endotherms. Proceedings of the National Academy of Sciences, 106(Supplement 2), 19666-19672.
Prothero, D. R. (2016). The Princeton field guide to prehistoric mammals (Vol. 112). Princeton University Press.
Rowe, M. F., Bakken, G. S., Ratliff, J. J., & Langman, V. A. (2013). Heat storage in Asian elephants during submaximal exercise: behavioral regulation of thermoregulatory constraints on activity in endothermic gigantotherms. Journal of Experimental Biology, 216(10), 1774-1785.
Špinar, Z. V., & Burian, Z. (1972). Life before man. McGraw-Hill Companies.
van Beest, F. M., & Milner, J. M. (2013). Behavioural responses to thermal conditions affect seasonal mass change in a heat-sensitive northern ungulate. PloS one, 8(6), e65972.
Weissenböck, N. M., Arnold, W., & Ruf, T. (2012). Taking the heat: thermoregulation in Asian elephants under different climatic conditions. Journal of Comparative Physiology B, 182(2), 311-319.
Wright, P. G., & Luck, C. P. (1984). Do elephants need to sweat?. South African Journal of Zoology, 19(4), 270-274.
Megaloceros giganteus galloping about: a rare scene in palaeoart, but probably a common sight in life. Famed for its size and headgear, we mostly ignore the fact that Megaloceros was among the most cursorial of all deer. What else about this amazing animal is hidden behind those antlers?
Megaloceros giganteus, otherwise known as the Irish elk or giant deer, can genuinely be regarded as a game-changing species for the study of extinct life. First discovered in 1695, it was a founding taxon of palaeontological science - a species that sent early scientists a clear message about the existence of life before the age of humanity and the reality of extinction. The significance of the Irish Elk - which, of course, is not strictly Irish nor anything to do with any living 'elk' species - was perhaps first fully grasped by Baron Georges Cuvier, who published a series of studies on giant deer in the late 1700s and early 1800s. Since then, Megaloceros has been an unwavering presence in paleontological media of all kinds: the focus of numerous scientific studies, a# case study for animal evolution, and a frequent star of palaeoartworks and popular texts. From a historic perspective, Megaloceros has demonstrated strong cultural staying power.
Among the earliest life reconstructions of Megaloceros is this small illustration from an 1854 issue of the periodical Die Gartenlaube. Its presence may have been influenced by the Crystal Palace Megaloceroscovered in the same magazine earlier that year. Although a relatively unremarkable restoration in most respects, it shows the same regal pose that has stereotyped portrayals of Megaloceros for nearly two centuries. If you've drawn or sculpted Megaloceros at some point, chances are it was standing somewhat like this.
An upshot of Megaloceros always being 'around' is that it's easy to overlook some of the cooler aspects of its palaeobiology. Its size and impressive antlers have become the principle takehome about its existence, such that other facts about its anatomy and ecology are ignored or forgotten. Recently, I've had cause to dig into the palaeobiology and research history of Megalocerosand, wow, what an animal. Cervids are genuinely fascinating anyway: charismatic, complex and awesome animals which, behind their soft, doleful eyes, have a physiology and life history evolved in partnership with Michael Bay. And it's within this context of deer being awesome, unusual animals that we can view Megaloceros as a kind of "superdeer": a species that represents the known extreme of many trends in cervid evolution, and that should be known for much more than its size and antler spread.
Giant deer 101
M. giganteus is a well-known taxon represented by thousands of fossils and with a wide but latitudinally-restricted distribution through Pleistocene and Holocene deposits of Europe and Asia (Lister 1994; Geist 1999; Vislobokova 2012; Lister et al. 2019). Like many Ice Age species, its distribution was continually shrinking and expanding in response to glacial growth and it never occupied the full extent of its known range at any one time. As with the mastodons discussed here recently, M. giganteus was an Ice Age species that avoided the cold. Although an adaptable animal that seems to have lived in a range of environments, Megaloceros appears to have thrived mostly during interglacials when the well-vegetated, herbaceous habitats it preferred were at their broadest (Vislobokova 2012). It lived in largely open, flat and well-watered settings termed ‘boreal parkland’ or ‘boreal steppe-woodland’, which were situated between cold steppe-tundra in the north and warmer, arid regions to the south. This parkland habitat provided Megaloceros with a diversity of grazing and browsing opportunities thanks to the presence of pine and spruce trees, as well as dense shrubs and herbs. Theprecise habitat preferences of Megaloceros changed over time and is reflected in antler shape: early subspecies had smaller, more erect antlers and seems to have preferred slightly more forested settings, for example (Lister 1994). Access to good quality food and water were likely critical to Megaloceros life histories due to the nutritional demands of growing those huge antlers each year (Geist 1999), and their need for highly productive plant communities likely prevented them from attaining the wider geographic ranges of other famous Pleistocene megafauna (Vislobokova 2012).
Distribution of M. giganteus sites recently compiled by Lister and Stuart (2019). Note the relatively limited latitudinal range.
Staying put in food-rich environments is probably one reason Megaloceros became so large and well-ornamented. Although not actually the largest extinct deer, the biggest stags are estimated to have massed an impressive 550-600 kg (or more) and stood about 2 m at the shoulder. Their antlers were the biggest of any cervid, weighing 35-45 kg and spanning up to 3.5 m. Reinforced skull bones and mandibular mineral storage were just some of their cranial adaptations to housing these immense organs (Lister 1994). Our knowledge of Megalceros does is comparably poor compared to that of stags, as our giant deer record strongly skews towards male specimens. So far as can be ascertained from a handful of skulls - the only elements we can routinely refer to either sex - females were of generally similar size to males, albeit about 80% of their mass and lacking antlers (Geist 1999). It's thought that our male-dominated fossil record reflects stags foraging separately to females and seeking aquatic vegetation rich in nutrients for antler growth. This, in turn, brought them more routinely into settings conducive to fossilisation. Although some fossil localities contain dozens or hundreds of Megalceros fossils, they are generally rare outside of these exceptional sites. This may reflect genuinely low populations of Megaloceros in most ancient environments (Lister 1994; Geist 1999), perhaps as a consequence of their resource needs.
Discussions and depictions of Megaloceros are heavily biased towards male specimens but - guess what? There were females too. Their fossils are significantly rarer than those of males but seem to have been of similar linear dimensions, though they were perhaps more slender overall. This skeleton is on display in the Natural History Museum, London.
Since the mid-1800s scholars have discussed where Megaloceros fits within deer phylogeny. Although firmly placed within Cervinae - the Old World deer clade - opinions differ over which extant deer are its closest relatives: Cervus (e.g. red deer, wapiti, sika deer - Kuehn et al. 2005) or Dama (fallow deer - Lister 1994; Geist 1999; Lister et al. 2005; Hughes 2005). The weight of morphological and DNA evidence for the Dama link is probably heavier at present, although this only resolves part of our struggles: how Megaloceros is related to other extinct deer is also uncertain. The composition, and even existence of a Megaloceros-line clade, Megacerini, is currently debated, as is whether Dama is the last surviving megacerine (see Vislobokova 2012 for a review).
Extreme deer, extreme speed
Prehistoric animals tend to become popular because they represent some kind of biological extreme - the biggest, the heaviest and so on. It's curious, therefore, that the running capacity of Megaloceros is neither widely remarked upon nor commonly depicted in palaeoartworks. A wealth of anatomical data shows that Megaloceros was among the most cursorial of all deer - a species adapted for running far and long over wide, open ground (Geist 1999; Janis et al. 2012). Deer exhibit a range of running styles but Megaloceros seems to have been most suited to long-distance galloping with low limb ground clearance, in contrast to species which bound at height over rough terrain or high-step at speed through dense vegetation. Reindeer and wapiti are similarly adapted to long-distance running, though neither is comparable in cursorial adaptation to Megaloceros. Along with cursorial limb proportions, Megaloceros also has an evident muscle bulk consistent with running habits, highly 'hinged' shoulders permitting an enhanced forelimb stride length, and an expanded trunk volume for an enlarged heart and lungs (Geist 1999). Relatively small hooves indicate a preference running over firm ground. I'm not aware of any specific estimates of Megaloceros running speed, but Geist (1999) considered Megaloceros comparable to the fastest living deer - perhaps 80 kph (50 mph) - and assumed similarly commendable levels of stamina. The increased bulk of Megaloceros would have made for slower acceleration but, once at cruising speed, it might have been capable of running for hours and hours without stopping.
Many aspects of Megaloceros osteology - from limb proportions to chest volume, antler configuration and knee morphology - are consistent with a strongly cursorial lifestyle. It's ironic that this energetic, fast-running deer is often depicted in static poses in palaeoart when dynamic and active compositions typify the genre as a whole. Image by James St. John, from Wikimedia Commons, CC-BY 2.0.
For Geist (1999), a running lifestyle is integral to explaining the development of Megaloceros size and antler spread. It stands to reason that the largest deer will also be the fastest and fittest, and antler size is a direct, honest reflection of stag health. These factors alone could have promoted the evolution of enormous antlers in a large cervid living in uncluttered habitats, but cursoriality encourages antler size in other ways too. For mammals, a consequence of evolving fast running is that juveniles have to be strong and coordinated enough to keep up with their parents, especially if running is critical to escaping danger. Megaloceros fawns must thus have also been capable runners from a very early age, requiring their parents to not only give birth to highly developed, precocial offspring but also supply them with milk rich enough to sustain long-distance, high-velocity running. Lo and behold, studies show that antler size correlates not only with offspring health and size, but also the amount of fat and nutrients in milk (Geist 1999). Put together, these factors mean Megaloceros wasn't a fast-running deer that happened to have huge antlers: its antlers were a direct consequence of its hyper-cursorial lifestyle.
But seriously though, those antlers
As is already evident, it's difficult to write about Megaloceros without frequent mention of its headgear, so let's tackle that topic head-on. There's a lot more to talk about here than just size. Antler structure, function and history of interpretation are also fascinating. Megaloceros antlers are extremely long - up to 1.7 m each - and broaden into great palms in their distal regions, with huge tines erupting from the burr, beam and palm margins. The first tine, which emerges just above the antler base, is broadened to differing extents in different subspecies, and is especially large in early representatives of the lineage (Lister 1994). This likely served as some sort of eye protector during combat (see below).
The size and elaboration of M. giganteus antlers placed it at the heart of a historic discussion about the fitness, evolution and extinction of fossil animals. M. giganteus stags were considered examples par excellence of orthogenic evolution in the late 19th and early 20th century when, as reviewed by Gould (1974), it was a species that had evolved itself into a corner: the result of a runaway, one-way evolutionary process that encouraged the creation of vastly oversized and biologically untenable antlers. Eventually, it was suggested, the antlers became so large and heavy that their owners were forever being caught in vegetation, mired in mud and bogs, or even suffered catastrophic brain haemorrhages caused by redirected blood flow from antler velvet. Extinction was inevitable for such sorry creatures.
Of course, even an elementary grasp of modern principles of natural selection shows these ideas as naive, quaint and totally wrong - there is no conceivable mechanism through which species can evolve structures that are so hazardous to their health. But it wasn't until the 1930s that a sensible alternative explanation for Megaloceros antler size was proposed (positive allometry), and we waited until the 1970s for someone to actually test whether Megaloceros antlers were actually oversized, cumbersome organs or consistent in proportion to what we'd expect from living deer. This study came in the form of Stephen J. Gould's famous 1974 paper "The origin and function of 'bizarre' structures: antler size and skull size in the 'Irish Elk,'Megaloceros giganteus", which demonstrated that Megaloceros antler dimensions were not only consistent with those predicted from living deer, but that their evolution was explainable through regular old sexual selection - no crazy, uncontrolled evolutionary mechanism required. Further work has shown that, relative to body size, Megaloceros antlers were no heavier than those of fallow deer and, indeed, were actually lighter than those of reindeer (Geist 1999). Gould's results were an important grounding of Megaloceros in the wider context of deer biology. Rather than seeing it as a weird, dead-end outlier, Gould showed that Megaloceros represented the known limit of antlers operating as 'honest' signals of stag health and virility. Their size is a predictable outcome of cervine evolution should species develop into large-bodied, open-country specialists.
Were Megaloceros antlers exceptionally large? In an absolute sense, yes, but in a relative sense, no: they're appropriate for a deer of its body size. This graph from Gould's landmark 1974 paper is a little crude - neither shoulder height nor antler length are the best measure of body proportions (masses would be preferable) but this study marked the beginning of more informed and quantified investigations into the evolution, function and significance of giant deer antlers.
Furthermore, ideas that Megaloceros antlers were somehow evolving out of control ignores nuances of their structure and evolution. There's no doubt that - as with all stags - giant deer males were physiologically stressed by antler production. Even assuming development occurred over several months, centimetres of growth were required each day and it's doubted that enough calcium and phosphate could be processed directly from their food fast enough to meet demand (Moen et al. 1999). Megaloceros responded with the standard cervid adaptation of borrowing bony material from its skeleton, and physiological models suggest up to 80% of their available resorbable calcium was required to achieve a full antler spread - about twice that of moose (Moen et al. 1999). Assuming access to suitable plants, these resorbed minerals could be replaced in time to rebuild a strong, durable skeleton for the rut, but osteoporosis remained a risk for stags unable to recapture that material. It's thus unsurprising that Megaloceros antlers are, on average, not quite as solid as those of other deer. All antlers are a blend of compact bone (very dense and heavy, equating to a high mineral cost) and a spongier component (lighter and less dense, so lower mineral cost), and Megaloceros evolved a higher ratio of lower-density bone to better manage the physiological demands of antler growth. Even this was not enough to prevent their antlers from becoming a burden at times, however: average antler size is seen to reduce on occasion throughout their evolutionary history. These responses are the exact opposite of what we'd expect in an orthogenic 'runaway evolution' model, and are evidence of how ordinary selection pressures were keeping Megaloceros anatomy in step with environmental conditions.
Megaloceros antlers were exceptional communication devices, appearing enormous from multiple angles, and especially so from the front. Unlike other deer, Megaloceros would not have to pose to show off its antler palms: they were unmissable however you saw it. Screengrab of a 3D scan by National Museums Scotland that is loads of fun to play with (and forms a useful reference for artists!).
The relatively low mineral value and size of Megaloceros antlers has led to discussions about their function: were they used for visual communication only? Palmate antler regions, which occur today in species like fallow deer and moose, are thought to serve primarily in communication, and it stands to reason that this applied to Megaloceros too. But while living palmate stags use head and neck motion to draw attention to their antlers (Gould 1974; Geist 1999), Megaloceros headgear was rotated in such a way that stags could exhibit their huge palms by simply standing still. This precluded the need to wave 40 kg of bone around for days on end, which was probably terrific news for the neck and head tissues of Megaloceros stags (Gould 1974). This said, Megaloceros skulls, necks and shoulders were heavily augmented to support their antlers (the skull roof was 30 mm thick! - Lister 1994) and it's probable that they were capable of moving them around with speed and precision, perhaps even violently. Although some authors (Gould 1974) have regarded Megaloceros stags as incapable of wrestling with one another, studies of their antler histology and stress distribution suggest such matches were possible (Lister 1994; Klinkhamer et al. 2019). The effects of clashing antlers have yet to be modelled - this being fraught with variables that are difficult to estimate at present - but Megaloceros antlers perform well under pushing and twisting regimes provided the stresses are primarily experienced in their proximal regions. They appear to have been especially resistant to rotational forces, and we might imagine stags locking their antlers together at close range, faces low to the ground, shoving and twisting each other to the floor (Lister 1994). Their antlers are so long that forces incurred at their tips might have been amplified to potentially bone-snapping levels however, so it's possible Megaloceros may have avoided especially violent, unpredictable fighting styles. Again, this is consistent with trends seen across deer: as stags become larger, and species become more sociable, their fights generally become more ritualised and lower risk. Megaloceros may be the ultimate example of this correlation (Geist 1999).
Although no soft-tissue Megaloceros fossils are known, a reasonable amount has been inferred about its life appearance through predictions of ecology and interpretation of Palaeolithic art(e.g. Lister 1994; Geist 1999; Guthrie 2006; also see Darren Naish's summary at Tetrapod Zoology). Although these differ in some details, a consensus has formed around some aspects, including the presence of a large shoulder hump (a diagnostic feature of Megaloceros cave art) and a fallow deer-like prominent laryngeal region. These features seem common to both males and females, implying a use beyond male competition and sexual signalling. It's tempting to link large larynxes with deep, guttural calling, akin to that produced by Dama, and with both genders sporting them, perhaps Megaloceros was an especially vociferous deer? Palaeolithic artists depicted the shoulder hump as large and bulging in both genders, so it probably does not merely represent the augmented, antler-carrying shoulder and neck muscles of stags. Geist (1999) speculated that it represents a fatty hump: an energy reserve for periods of seasonal stress that would not, by being concentrated in one place, cause the same overheating risk as distributing fatty tissues across the body, as occurs in some reindeer. Zebu cattle, white rhinos, bison and buffalo are similarly adapted running species (Geist 1999). There is no artistic evidence of a mane or bell, as seen in some other large cervids, although some Palaeolithic art hints at longer hairs on the hump.
A take on Megaloceros body patterning based on a very literalistic reading of certain Palaeolithic artworks - note the stripe running from the shoulder to hindlimb on the stag. The shoulder hump on the doe should probably be larger.
Predictions about colouration have also been presented, although no-one should be under the illusion that we have a robust idea of colours and patterns in this species. Our most informative insights into this are cave art showing dark colours at the shoulder hump which taper into dark stripes extending across the body and neck (diagonally across the flank from the shoulder to the knee region, as well as vertically across the neck-torso junction). A further dark stripe or band is recorded at the top of the neck, possibly in relation to the enlarged laryngeal region. Another vertical stripe may have framed a rump patch - a feature consistent with Dama. How we interpret these drawings isn't 100% clear, with the full-body restorations of Geist (1999) and Guthrie (2006) differing in some details. Geist regarded Megaloceros as pale all over with a conspicuously dark transverse body stripe and dorsal midline, while Guthrie portrayed more regionalised colouration with darker hindquarters grading into paler hues around the shoulder, framed by prominent stripes. Lister (1994) was more conservative, merely noting the likelihood of the dark shoulder hump.
One of the most informative Palaeolithic artworks of Megaloceros is featured in the Megaloceros Gallery in France's famous Chauvet Cave. Unlike other examples, it seems to show shaded areas that could reflect darker body regions, similar to the banded Coelodonta depicted on the opposite wall. A second Megaloceros is featured a little further in, also showing a dark neck band. Alas, other Megaloceros panel drawings lack similar details, precluding determination of their reliability. Cropped from Google Arts and Culture - make sure to check out the Megaloceros Gallery in 3D!
The challenges of interpreting Megaloceros colouration from cave art alone are many. Opinions differ on how many panel drawings we have - perhaps up to 40, but of which maybe only 15 or 16 are confidently identified as M. giganteus. Most are body outlines without details of colours or patterns and, to my knowledge, only 4-5 pieces feature those dark humps and body stripes. This is enough to assume that they are recording something about Megaloceros life appearance, but what, exactly, might that be? Are the lines radiating from the shoulder humps stripes, or something else, like the boundaries between colour regions? Are those simpler 'body outline' artworks actually depicting Megaloceros with uniform colouration? Do these differences reflect seasonal or regional colour variation, or else varying artistic approaches? The question of how literally we should take Palaeolithic depictions is not new, and it doesn't have a straightforward answer (Guthrie 2006).
Select Palaeolithic art of Megaloceros as interpreted by Guthrie (2006) - arrows indicate features of interest such as shoulder humps, expanded laryngeal regions etc. Note the absence of obvious colour and pattern data in most depictions - what significance, if any, might that have? This compilation of Guthrie's work is swiped from Tetrapod Zoology.
But while Palaeolithic art does not give us a clear answer on Megaloceros life appearance, ideas of it being a highly-specialised 'extreme' deer species help us explore likely colour schemes a little further. We can look to deer and other ungulates that match Megaloceros in behaviour and ecology to make some broad-brush predictions of what an especially large, well-ornamented, communicative, open-habitat and cursorial deer species may have looked like. Geist (1999) calculated that an energetic 600 kg deer living in an open, shadeless environment could be sensitive to solar insolation, especially during sustained ruts and runs, and predicted light-colours across the body to reflect sunlight and avoid overheating. Thoughts that Megaloceros may have been subject to heat stress are not baseless: some readers may recall that moose are highly prone to overheating, even in winter, despite living in shadier settings and having a higher surface area:volume ratio than Megaloceros (Dussault et al. 2004; van Beest and Milner 2013). Geist also predicted a rump patch - a feature seen in Dama as well as possibly one piece of ancient art - as this feature has convergently developed in a number of social, cursorial ungulates. The possibility of dark body regions or stripes has some functional explanation, too, with Geist (1999), Guthrie (2006) and Lister (1994) all noting the utility of dark patterning in ornamented deer, and Geist suggesting this would be especially effective in an open environment against a light sky. It's difficult to know how correct these predictions are of course, but I admire the fact that they consider the unique lifestyle of Megaloceros rather than, as has been palaeoart tradition until recently, merely dressing Megaloceros as a red deer or wapiti.
Charles Knight's depiction of a Megaloceros stag being hunted by early humans. Did we contribute to the extinction of giant deer? Possibly, although evidence for our interaction with this species is uncommon. From Knight (1949).
It seems remiss to have written all this about Megaloceros without mentioning modern ideas of why it is no longer with us. This remains a topic of ongoing research, and a synthesis provided by Lister and Stewart (2019) gives a good overview of the current state of the debate. Our vast sample size and detailed understanding of Quaternary sediments means that the history of Megaloceros can be read in detail, providing a substantial (though still developing) insight into the major factors that contributed to its demise. Once thought to have been an exclusively Pleistocene species that held out longest in northwest Europe, recent discoveries and dating of Megaloceros sites in Siberia have not only confirmed its survival well into the Holocene, but shown that Asia held Megaloceros populations longer than anywhere else - as recently as 7,600 years ago.
With concepts that Megaloceros antlers were some sort of doomed anatomy pulling giant deer to extinction firmly disregarded, more sensible discussions about Megaloceros demise consider environmental change, their high productivity demands, and human hunting likely extinction causes. Generally, it seems that local Megaloceros extinctions coincide with habitats changing from parkland habitat to wooded or tundra-like settings, both of which heralded reduction in optimal foraging conditions as well as losses of plants rich in calcium. Megaloceros populations in those areas may have thus dwindled or been forced to find more suitable places to live. There is evidence that Megaloceros anatomy responded somewhat to nutrient-stressed conditions, but ideas that dwarfed or otherwise stunted populations developed in islands or other refugia have not been bourne out: late-surviving Megaloceros were only something like 2% smaller than earlier specimens. The lack of more dramatic body size reduction may be related to predation pressures to stay large and fast, especially as new data shows that humans co-existed with Megaloceros for longer than was once thought. Evidence for our hunting giant deer is relatively rare so we may not have been a major factor in their extinction, but as suitable habitats reduced, even low hunting frequencies may have been enough to push Megaloceros over the edge.
As is always the case with a well-studied species, there's much more we could say about Megaloceros. We are still learning about its palaeobiogeography, behaviour, evolution and extinction from new discoveries and detailed examination of archived specimens. Digging into the literature on these animals has proven genuinely fascinating. As someone used to studying Mesozoic reptiles, the amount of information and insight we have on this species is almost overwhelming, and it seems that understanding its palaeobiology is challenged more by having to consider so much data rather than, as I'm more accustomed to, having too little information to draw meaningful conclusions. And yet, despite an excellent fossil record and centuries of study, some major aspects of its biology remain poorly understood or contested. Food for thought, indeed, about our capacity to interpret fossil tetrapods from more typical palaeontological datasets.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Dussault, C., Ouellet, J. P., Courtois, R., Huot, J., Breton, L., & Larochelle, J. (2004). Behavioural responses of moose to thermal conditions in the boreal forest. Ecoscience, 11(3), 321-328.
Geist, V. (1999). Deer of the World. Swan Hill Press, Shrewsbury.
Gould, S. J. (1974). The origin and function of 'bizarre' structures: antler size and skull size in the 'Irish Elk,'Megaloceros giganteus. Evolution, 191-220.
Guthrie, R. D. (2005). The Nature of Paleolithic Art. The University of Chicago Press.
Hughes, S., Hayden, T. J., Douady, C. J., Tougard, C., Germonpré, M., Stuart, A., ... & Say, L. (2006). Molecular phylogeny of the extinct giant deer, Megaloceros giganteus. Molecular phylogenetics and evolution, 40(1), 285-291.
Janis, C. M., Shoshitaishvili, B., Kambic, R., & Figueirido, B. (2012). On their knees: distal femur asymmetry in ungulates and its relationship to body size and locomotion. Journal of Vertebrate Paleontology, 32(2), 433-445.
Klinkhamer, A. J., Woodley, N., Neenan, J. M., Parr, W. C., Clausen, P., Sánchez-Villagra, M. R., ... & Wroe, S. (2019). Head to head: the case for fighting behaviour in Megaloceros giganteus using finite-element analysis. Proceedings of the Royal Society B, 286(1912), 20191873.
Knight, C. R. (1949). Prehistoric man: The great adventurer. Appleton-Century-Crofts.
Kuehn, R., Ludt, C. J., Schroeder, W., & Rottmann, O. (2005). Molecular phylogeny of Megaloceros giganteus—the giant deer or just a giant red deer?. Zoological Science, 22(9), 1031-1044.
Lister, A. M. (1994). The evolution of the giant deer, Megaloceros giganteus (Blumenbach). Zoological Journal of the Linnean Society, 112(1-2), 65-100.
Lister, A. M., Edwards, C. J., Nock, D. A. W., Bunce, M., Van Pijlen, I. A., Bradley, D. G., ... & Barnes, I. (2005). The phylogenetic position of the ‘giant deer’ Megaloceros giganteus. Nature, 438(7069), 850-853.
Lister, A. M., & Stuart, A. J. (2019). The extinction of the giant deer Megaloceros giganteus (Blumenbach): New radiocarbon evidence. Quaternary International, 500, 185-203.
Moen, R. A., Pastor, J., & Cohen, Y. (1999). Antler growth and extinction of Irish elk. Evolutionary Ecology Research, 1(2), 235-249.
van Beest, F. M., & Milner, J. M. (2013). Behavioural responses to thermal conditions affect seasonal mass change in a heat-sensitive northern ungulate. PloS one, 8(6), e65972.
Vislobokova, I. A. (2012). Giant deer: origin, evolution, role in the biosphere. Paleontological Journal, 46(7), 643-775.
Dinosaurs and dragons: is there a better fit between fossils and folklore? Common wisdom is that Chinese dinosaur species, perhaps including Yutyrannus huali, became associated with local dragon lore when ancient people used their bones for medicinal purposes, but how well-evidenced is this popular idea?
Geomythology is a discipline that most of us are familiar with even if we've not heard of this term: the study of possible associations between real geological phenomena and myths and legends. The idea that certain fossils were somehow involved in the creation or development of mythical creatures is a subject we've discussed several times at this blog, including the purported fossil associations of griffins, cyclopes, giants and unicorns. Although proverbial kernels of truth underlie some of these proposals, many examples - including famous claims about Protoceratops and griffins, and elephant skulls and cyclopes, are actually nowhere near as well-supported as their popularity implies. A lot of geomythological hypotheses persist primarily because of uncritical retellings and a lack of sceptical examination.
Today, we're going to take aim at another claimed geomythological association: the proposal that fossils of Mesozoic dinosaurs were mistaken for dragon bones in ancient and historic China. This idea only has a slim footing in academic literature but is very popular, being brought up just about whenever the early history of dinosaur discoveries is raised and being bolstered by mentions in reputable outlets like Smithsonian Magazineand BBC Earth. A common issue with arguments in favour of this hypothesis, whether written in a technical paper or posted to Youtube, is vagueness: beyond pointing out the antiquity of fossil discoveries in China and the ancient origin of dragon myths, little more is said. But we don't need to be vague: we actually have extensive documentation of historic 'dragon bones' in the form of numerous historical texts, illustrations, and records of interaction between 19th and 20th-century scientists with the dragon bone industry. Collectively, these create excellent insights into what 'dragon bones' really were and what fossil animals they belonged to. So how does the dinosaur-dragon association stand up against this evidence?
Cropped dragon from the 17th-century painting Miracle of the Dragon. Chinese dragon lore shares many features with dragon myths in other Asian cultures, so they likely have a principally folkloric, not palaeontological, origin. You can see the entire and spectacular Miracle of the Dragon painting at the Cleveland Museum of Art website.
Dragon mythology has existed in China since at least the Shang Dynasty (1520 - 1030 BCE) - in other words, for as long as we've had decent records. It likely originated among the myths of other cultures rather than being a novel creation based on fossils: Hogarth (1979) suggests that the older Buddist naga dragon myth has so many cultural similarities with Chinese dragon lore that the two stories must be linked. This said, ancient texts also show that Chinese scholars knew of fossil bones thousands of years ago, and it's primarily these documents that are used to make the case for the dinosaur-dragon association (e.g. Needham 1959; Dong 1988; Sarjeant 1999; Spalding and Sarjeant 2012; Rieppel 2019). Dong (1988) and Rieppel (2019), for example, suggest that scholar Qu Chang may have been discussing dinosaur fossil beds in what is now Sichuan Province in c.350 CE. Rieppel (2019, p. 236) provides a direct translation:
"In Wucheng Countym there is a mountain called Somber Warrier Mountain, also called Three Corner Mountain, that has six bends and six rises. Dragon bones are taken from it. It is said that dragons flew up from these mountains, but when they found heaven's gates closed, they could not enter, and thus fell dead in that place, and later sank into the earth. That is why one can dig out dragon bones."
Dong (1988) further notes that Jurassic strata in this area may be the real-life source of Chang's 'dragon bones', which seems a reasonable prima facie argument.
Another ancient account, cited in some popular articles, recalls the recovery of a 'dragon bone' during the construction of a canal in 120 BCE. This discovery was so significant that the waterway was named "Dragons Head Canal" after its discovery (McCormick and Parascandola 1981). Several other records of such antiquity are known and make a fine case for ancient Chinese cultures interacting with fossils, but they each have the same problem: none are detailed enough to demonstrate that they pertain specifically to dinosaur fossils (Delair and Sargeant 1975; Buffetaut 1987; Sarjeant 1999). Dinosaurs don't just occur anywhere and everywhere fossils are found, so without descriptions or illustrations of the bones, or specific geographic information we can use to track down these ancient sites, we can only guess what these first mentioned 'dragon bones' were. Qu Chang's discussion of the Sichuan Province is interesting because - as noted by Dong - this is a rich ground of dinosaur fossils, but the same province also bears plenty of non-dinosaur fossil sites, including fossil-rich Pleistocene gravels (Delair and Sarjeant 2002) and sinkhole deposits (Buffetaut 1987). Without more data, we simply cannot say what fossils these ancient documents pertain to, and the involvement of dinosaurs is equivocal.
An honest-to-goodness collection of 'dragon bone' teeth obtained from drugstores during the late 19th century, as figured by McCormick and Parascandola (1981). No doubt some readers are already identifying some of these elements: many are quite obvious to anyone generally familiar with vertebrate anatomy.
And that, really, is that. If you're hoping that our tale now turns to more concrete evidence of the Chinese dragon-dinosaur link, you're out of luck: these loose, arm-wavy arguments are the best we have to offer. The crux of our problem is that 'dragon bone' in ancient China was a catch-all term for a variety of fossil types that were initially only categorised as being bone, tooth or horn. By the 5th century, colour and texture were also factored to determine dragon gender and bone quality (McCormick and Parascandola 1981), but something resembling modern taxonomy was never applied. Economic value and pharmaceutical use were the main interest in 'dragon bones' so it was along these lines that they were categorised: details of anatomy and morphology were basically unimportant. We know this thanks to medical documents dating back as far as the third century BCE. Dragon remedies were considered powerful agents that could treat dysentery-like symptoms, heart, liver and kidney conditions, forgetfulness, anxiety, epilepsy and even disembowelment (McCormick and Parascandola 1981; Buffetaut 1987). You may have eaten dragon bone raw, boiled it, fried it, cooked it in rice wine, or else crushed it into powder to combine it with other ingredients. Ground bone sprinkled into tea eventually became the most common way of taking your dragon medicine (Hargarth 1979).
Alongside the medicinal use of dragon remains, the nature of the dragons themselves were also routinely discussed by Chinese physicians: principally, whether the dragons these bones were harvested from were alive or dead (some argued that dragons shed their bones like a snake sheds its skin). Little interest was shown in the geographic and geological origins of the fossils themselves, with some exceptions, such as a medical text from 456-536 CE that outlined dragon bones being extracted from caves along rivers (McCormick and Parascandola 1981). Although still vague, this helps us in our investigation to some extent by implying that some dragon bones were not especially old. Fossils collected from caves tend to be thousands of years old, not millions, so Mesozoic dinosaurs were unlikely to be among the fossils referenced in this text.
Dragon bones were just one of several fossil commodities available in historic China. As early as the 12th-century Chinese scholars had correctly identified that animal remains were capable of becoming petrified (Wen-chung 1956) and specimens of fossil shellfish and fish were commercially available for decorative and pharmaceutical purposes (Needham 1959; Edwards 1967). Fossil fish were so sought after that a market in forged specimens was established (Buffetaut 1987) and dragon bones were also in extremely high demand. This might be one reason why their source localities were not widely recorded - suppliers may have been cagey about their suppliers to protect their commercial interests. Contrary, perhaps, to Western expectations, dragon bones were big business for the Chinese pharmaceutical industry and were given premium prices (Hargarth 1978; Duffin 2017). They were sold in huge quantities from drugstores around the country and even exported en masse to neighbouring regions. Buffetaut (1987) mentions an 1885 manifest detailing some 20 tonnes of dragon bones passing through a Chinese port in a single year, and even in the 1980s several tonnes of 'dragon bones' were being exported internationally. At one time Chinese dragon bones were available in Chinese communities across Japan, Tibet, the East Indies, the Philippines, and the USA (Koenigswald 1952; McCormick and Parascandola 1981). And yes, you can still walk into some Chinese pharmacies and buy dragon bone today.
These qualities also have bearing on our investigation of dragon bone origins. Their widespread availability and mass-exportation tell us that they were not difficult fossils to find, extract or transport. Rather, they must have been superabundant, robust fossils that could be easily dug and cleaned using the basic technology of historic rural China (Wang et al. 2020). If so, this conclusion also doesn't help the dragon-dinosaur hypothesis. Mesozoic dinosaur bones can be abundant but, on account of their age and the lithification of the sediments they occur in, they are often challenging to excavate and can be very fragile. Most of us will be familiar with the careful excavation, jacketing and preparation processes that accompany modern dinosaur discoveries - it's unlikely this sort of time-consuming care was practised thousands or hundreds of years ago in the Chinese countryside.
1505 illustration of 'dragon bones' from the Bencao pinhui jingyao, more recently featured by Duffin (2013). This illustration is significant for showing us what constituted a 'dragon bone' some 500 years ago, several centuries before scientists saw and began documenting these remains.
It's through China's traditional medicine industry that 19th-century European scholars first documented China's vertebrate fossils. It was this process that transferred dragon bone mythology into scientific reality. Europeans visiting China bought and sent 'dragon bone' specimens to their home institutions at a grand scale, cumulating in thousands of bones and teeth being sent to Western museums (Buffetaut 1987). This material was exciting to 19th-century scientists and attracted the attention of premiere scholars, including Richard Owen, who provided the first scientific description of Chinese vertebrate fossils in 1858 (see Owen 1870) and returned to this topic several times in his career. What Owen and others discovered was that Chinese 'dragon bones' was a catch-all term for fossil debris: shells, small bones, bone fragments, teeth and horns. Some bones had evidently been more complete when found but were broken to be sold, especially if it meant relatively valuable teeth could be freed from the less desirable jawbones (Koenigswald 1952). Some bones retained traces of their source sediment but exact locality data were unrecorded.
Despite the lack of provenance and quality of the fossils, European and (later) American palaeontologists were able to identify numerous species among the dragon bone collections. The Devonian brachiopod Spirifer verneuili was among the most common elements sent to European collections - a fossil also thought to have curative properties in traditional Chinese medicine - but vertebrate material was also present in abundance: specifically, Neogene and Pleistocene mammals. By far the most common vertebrate elements were the teeth of the Miocene-Pleistocene equine Hipparion, of which thousands of examples were sent to Europe alone (Koenigswald 1952; Buffetaut et al. 1987). Many other species were present too: some 60 mammal species were identified in one large sample analysed in Munich (Koenigswald 1952). Over time, 'dragon bones' from across China were found to include fossil horses, giraffes, elephants, rhinos, tapirs, hyenas, deer (their antlers being 'dragon horns'), bears and hominids (Koenigswald 1952; McCormick and Parascandola 1981; Buffetaut 1987; Duffin 2008, 2013; Wang et al. 2020). Among the most exciting finds to come from the drugstore dragon bones were hominid remains, which helped catalyse interest in Asia as a source of human ancestry. The discovery of Peking Man can be directly related to 'dragon bones' and the first known fossils of everyone's favourite enigmatic giant fossil ape, Gigantopithecus, were purchased over a drugstore counter (Koenigswald 1952; Wen-chung 1956; McCormick and Parascandola 1981). That mammal bones were being sold as dragon remains for hundreds of years before European scientists became interested is documented by early 16th-century illustrations of 'dragon bones' which are unmistakably mammal teeth and deer antlers (above, Duffin 2013).
The anatomical reality of Hipparion, a mid-sized, three-toed Miocene-Pleistocene horse, might seem millions of miles away from that of a Chinese dragon, but the teeth of this genus constituted the overwhelming majority of 'dragon teeth' reviewed by 19th and 20th-century scientists. Strange as it seems, the fossils of this horse have far greater relevance to Chinese dragon mythology than any Mesozoic dinosaur.
Excited by the flow of fossils from China but frustrated by their quality and lack of geological context, Western explorers and palaeontologists eventually gained access to dragon bone sites through the same networks that supplied Chinese drugstores (Koenigswald 1952; Buffetaut 1987; Wang et al. 2020). Among these were riverside caves in Yunnan Province and sinkholes in Sichuan Province, both of which were richly stocked with Pleistocene mammal fossils. Given that both riverside caves and Sichuan Province were mentioned by early accounts of fossil discovery in China, accessing these sites goes some way to closing the loop in our dragon bone story, although whether they are the exact same sites referenced in these ancient writings will forever be unknown.
One of the most detailed accounts of a dragon bone locality was provided by Walter Granger in his visit to a series of Pleistocene sinkholes in Sichuan Province during the 1920s. These contained abundant, well-preserved Pleistocene mammals that were excavated by local farmers lowering themselves into pits and digging fossils from soft mud. Once hauled to the surface, the fossils were cleaned with water before being stacked in rough piles to dry (Buffetaut 1987). Granger specifically noted that the fossils were treated roughly, there being no need to worry about their condition given their pharmaceutical destiny (Koenigswald 1952). Granger's description of recent-ish fossils being extracted from loose sediments is exactly the sort of low-tech set-up we'd expect to be supplying the dragon bone industry, and similar sites were reported by other explorers and geologists. Surprisingly, there are still dragon bone sites waiting to be found: some fossils brought back from China in the 19th and 20th centuries have yet to be matched to their source rocks. This is an active area of research as many of those exported bones are now type specimens that should, ideally, have established provenance (Wang et al. 2020) - a cautionary tale par excellence for the importance of labelling your specimens!
But wait wait wait: this is an essay about dinosaurs and dragons, and I'm waxing on about mammals. It's here, facing centuries of excellent documentation about the reality of 'dragon bones', that the lack of a substantiated link between dinosaurs and Chinese dragons is especially obvious. To my knowledge, no dinosaur bones were identified among the dragon bone samples sent to Europe and North America during the 19th and early 20th centuries, no illustrations of obviously dinosaurian fossils are known among historic works, and no dinosaur localities were discovered through drugstore dragon bone networks. Instead, China's dinosaur sites were made known to science through traditional geological prospecting and amateur discoveries in the early 20th century (Buffetaut 1987) - no dragons required. In contrast, we know for a fact that Chinese dragons were associated with extinct mammals, many of which are relatively familiar types - horses, hyenas, bears and so on. It's strange to juxtapose these animals with the fantastic dragons of Chinese mythology, but that's what our evidence shows.
Part of the surprising pervasiveness of the Chinese dinosaur-dragon link is that the real history of dragon bones is already associated with exciting, popular fossil species, including Homo erectus and Gigantopithecus blacki. In this light, it's hard not to see the perpetuation of the speculated, unsupported dinosaur-dragon history as overriding the genuinely interesting and significant contributions that Chinese 'dragon bones' have made to palaeontological research.
Of course, if we're being true sceptics on this, we also have to point out some important caveats. Firstly, our historic record of dragon bones is good, but it does not cover the entirety of China's geographic, historic and paleontological resources: we cannot rule out any and all use of dinosaur bone as dragon remains in historic times. It would actually be surprising, given the appetite for dragon bone and the abundance of dinosaur material in China, if no-one found and interpreted a dinosaur or another type of fossil reptile in this way. So who knows - evidence of this may turn up in future.
Secondly, although the historic use of dinosaur bones in traditional medicine is not well-evidenced, a surprise discovery in 2007 revealed that a central Chinese village has been using dinosaur bones in medicine for at least a few decades. It differs from older historic accounts in that these villagers have been collecting their fossils locally and not through drugstores, and makes me wonder if other small communities exploited dinosaur fossils in the same way. Maybe our records and analyses are biased towards the history of the national, commercialised end of dragon bone use? It's an interesting idea, but is also currently an unknown.
But these points don't change the details outlined above. I wonder if some dinosaur fans will be disappointed by the lack of evidence for a dinosaur-dragon link, but I think it's a fascinating story with a terrific twist: the fantastic and iconic Chinese dragon is most closely associated, palaeontologically speaking, with familiar mammalian megafauna - including our own lineage. But beyond providing a fun historic narrative, this outcome also serves as an example of how geomythological ideas can be misguided by our own biases and wishful thinking. I suspect we inject dinosaurs into this story because we see their obvious similarity with ancient dragon depictions, but this assumption falls foul of historian's fallacy: the projection of contemporary knowledge onto people of the past. Opinions vary on how knowledgeable ancient people were of dinosaur anatomy (Lyons 2009) but we have no evidence that the appearance of Mesozoic dinosaurs was known to anyone before complete dinosaur skeletons were discovered, excavated, prepared and reassembled in late 19th century Europe. My suspicion is that ancient knowledge of dinosaurs was no greater than what could be gleaned from partially-exposed, weathered fossils or loose bones and teeth: the alternative, that people living thousands of years ago in China had - for example - concepts of what whole stegosaurs looked like is very far-fetched. Moreover, our better-evidenced examples of geomythology, which includes Chinese dragons, shows that there need not be any real resemblance between fossil species and the legendary animals they become associated with: an ammonite can be a snake, a mammoth can be a unicorn, and a fossil horse can be a dragon. These might seem ridiculous to us, but only because centuries of scientific insight makes interpreting fossils and ordering the natural world look so easy. Our modern learned interpretations of fossils were not necessarily shared with peoples living thousands or hundreds of years ago.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Buffetaut, E. (1987). A short history of vertebrate paleontology. Croom Helm.
Delair, J. B., & Sarjeant, W. A. (1975). The earliest discoveries of dinosaurs. Isis, 66(1), 5-25.
Delair, J. B., & Sarjeant, W. A. (2002). The earliest discoveries of dinosaurs: the records re-examined. Proceedings of the Geologists' Association, 113(3), 185-197.
Dong, Z. (1988). Dinosaurs from China. British Museum of Natural History and China Ocean Press.
Duffin, C. J. (2008). Fossils and folklore. Ethical Record, 113(3), 17-21.
Duffin, C. J. (2013). Lithotherapeutical research sources from antiquity to the mid-eighteenth century. Geological Society, London, Special Publications, 375(1), 7-43.
Edwards, W. N. (1967). The early history of palaeontology (No. 658). Rudolph William Sabbott Natural History Books.
Hogarth, P. 1978. Dragons. Allen Lane.
von Koenigswald, G. H. R. (1952). Gigantopithecus blacki von Koenigswald, a giant fossil hominoid from the Pleistocene of southern China. Anthropological papers of the AMNH; v. 43, pt. 4.
McCormick, J. P., & Parascandola, J. (1981). Dragon bones and drugstores: the interaction of pharmacy and paleontology in the search for early man in China. Pharmacy in history, 23(2), 55-70.
Needham, J. 1959. Science and civilisation in China. Volume 3. Mathematics and the sciences of the heavens and the Earth. Cambridge University Press.
Owen, R. (1870). On fossil remains of mammals found in China. Quarterly Journal of the Geological Society, 26(1-2), 417-436.
Rieppel, L. (2019). Assembling the dinosaur: Fossil hunters, tycoons, and the making of a spectacle. Harvard University Press.
Sarjeant, W. A. 1999. The earliest discoveries. In: Farlow, J. O., & Brett-Surman, M. K. (Eds.). (1999). The complete dinosaur. Indiana University Press.
Spalding, D. A., Sarjeant, W. A., & Brett-Surman, M. K. (2012). Dinosaurs: the earliest discoveries. In: Brett-Surman, MK, Holtz jr., T. R., Farlow, James O. (Eds.): The Complete Dinosaur. Indiana University Press.
Wang, X., Jukar, A. M., Tseng, Z. J., & Li, Q. (2020). Dragon bones from the heavens: European explorations and early palaeontology in Zanda Basin of Tibet, retracing type locality of Qurliqnoria hundesiensis and Hipparion (Plesiohipparion) zandaense. Historical Biology, 1-12.
Wen-chung, P. (1956). New material on man's origins. East and West, 7(3), 268-270.
After a long wait and much online discussion, the Mary Anning-inspired historic drama Ammonite is finally on general release. As goes the popularisation of palaeontology, Ammonite is something of a big deal: it’s the first film treatment of the iconic 19th-century fossil collector Mary Anning, a rare major feature to focus on genuine 19th-century palaeontology, and - in what might be another first - is a palaeontology-inspired film aimed squarely at adults. It has also, however, been controversial since its announcement for not focusing on traditional aspects of the Mary Anning story, such as her significance to the discovery of Mesozoic marine reptiles, her relationship with palaeontologists of the early 1800s and her tragically short, poverty-stricken existence. Instead, the film invents a narrative about an imagined romance between Anning and another historic figure, the geologist Charlotte Murchison. The appropriateness of this angle and what it means for Anning’s legacy has been the subject of much social media discussion, although the actual release of the film on premium streaming services has not, to my knowledge, generated the same level of debate. Curious to see if all our hopes, fears and general anticipation were worth our time, I recently checked out Ammonite and have come away with a mixed reaction. Is it a good film? I thought so. Is it a good ‘Mary Anning film’? I thought not. Are these answers mutually exclusive? It depends what you want from your Anning cinematic experience.
Let’s talk about the positives first. There is a lot I liked about Ammonite. It’s well-acted, well-directed, and delivers an outwardly strong reflection on sexism and classism in a strongly patriarchal Victorian society. The film is not subtle in its messaging: the opening shots show a maid scrubbing a museum floor being rudely pushed aside by bustling, suited men rushing an Anning-discovered Temnodontosaurus to its cabinet. It does, however, do a commendable job of showing, without preaching about, the contrast between rich and poor and the gulf in privilege that existed, and still exists, between men and women. Anning’s simple clothes, her empty, tired home and shop, as well as the sometimes bleak Lyme Regis coastline contrast well against the wealth, comfort and extravagance of richer characters and establishments. It provides a warts-and-all look at life on the poverty line in the 1840s where Anning and her elderly, poorly mother eat thin vegetable stews, illuminate their home with solitary candles, and several shots show their rough, tattered hands resulting from a lifetime of hard graft. Actors are not prettied up to look miraculously glamorous despite their lifestyle: there's an honest rawness to their appearance and costumes.
The tone of Ammonite is reflected with a sparse musical score and suitably bleak, although not lifeless, colour palette. Anning’s house is the core location of the film, but grubby British mudstone cliffs are its second home. I admit to finding these fossiliferous landscapes a refreshing sight for the setting of a palaeontological drama instead of, as is so often the case in palaeo media, vast deserts or badlands. The ever-present roaring waves and changeable weather of the southern UK coastline are excellently captured, and the cinematography manages to balance the grey colours of the Dorset coast and Anning’s home with stronger hues, especially the blues of Anning’s clothing, and the sea and sky. The film has a washed-out, slightly tired look that doesn’t feel forced, and perfectly suits the rest of the subject matter.
The first act features what most will expect of a Mary Anning picture, showing her looking for fossils in poor weather and clambering up slippery cliffs to excavate nodules containing ammonites. We are not explicitly told when the film is set, but the film’s version of Anning, played by Kate Winslet, imagines her in her later years - so presumably in the early 1840s. Portrayed as an embittered, middle-aged and experienced fossil collector with little time or interest in social graces, she also has a physical presence and resourceful quality entirely atypical for a 19th century female character. It's hard not to see some of this as making Anning the match for any man you'd care to put in her position. She's a woman of few words, wears trousers under her dresses, pees wherever she likes in the field (and then wipes her hands on her clothes), smokes hand-rolled cigarettes and deploys several harsh swears. Although outwardly a cold, embittered character with little patience for others, she is not unlikeable, and Winslet’s portrayal is genuinely excellent, her face showing an unspoken history of sadness, loneliness and world-weariness that needn’t be explained through dialogue. Her performance and frame have a stiffness that is at once both imposing and awkward, and much of what Anning thinks and feels is conveyed through forced stillness and suppressed reaction. It's a terrific performance that has unsurprisingly drawn much praise from critics.
As you’re no doubt inferring, Ammonite is not a breezy, lighthearted film. Nor, in contrast to virtually all other palaeo-inspired motion pictures I can think of, is it a family film. Ammonite has a ‘15’ rating in the UK (equivalent to an ‘R’ in the US), and for good reason: there’s male and female nudity, graphic sex and several strong swears. This is not the film to show your kids the Mary Anning story: it’s a slow, character-driven drama aimed at mature audiences. Although seemingly ruling out large chunks of its potential audience, for science communicators, this is a Good Thing. The last century of palaeo-related cinema can be largely boiled down to people running from animated dinosaurs, and Ammonite is going to draw attention from audiences who have no interest in this sort of thing. There is no shortage of child-friendly Mary Anning media out there, so it's great to have something that will draw the attention of older audiences.
But it’s also on the science communication front that Ammonite is going to prove most divisive. My take on films, TV shows and so on is that we have to rate them based on what the creators set out to do, not what we wanted them to do, and in this sense Ammonite might be free of criticism over its loose take on history. But I think it’s fair to ask whether Ammonite really needed to hang its narrative around Mary Anning at all, such are the liberties it takes with the subject matter. I have two main thoughts on this.
First, for a story about one of the most famous fossil hunters in history, Ammonite is strangely empty of what we might call palaeontological character. I understand that Ammonite is not a Marvel film or Star Trek episode where fans are deliberately fed blink-and-you-miss-them references, callbacks and easter-eggs, but the early 1800s yielded so many iconic specimens, books and palaeoartworks that I was surprised the film was so stripped back of palaeo-based content. Anning’s shop and home are virtually empty of specimens, which runs contrary to just about every fossil collector home, shop and lab I've ever been to. Invariably, such locations are full of stuff related to extracting and understanding extinct life: field gear, books, fossils, notes, rocks, unprepared specimens and other curios. I think the emptiness of her home is meant to stress her poverty, but the effect was that she looked more like she was a hobbyist fossil collector rather than the grandmother of palaeontology.
This bareness has another effect: it denies the sense that Anning made any multiple significant discoveries. We see Anning collect and prepare an ichthyosaur skull at one point but, other than this, a bookending cameo by the famous Temnodontosaurus fossil Anning found with her brother (the first ichthyosaur studied by scientists) and a quick shot of Anning sketching the Plesiosaurus dolichodeirus holotype from memory, there’s really nothing to represent her remarkable contribution to marine reptile palaeontology. It’s these discoveries that Anning is primarily remembered for and their absence will be noticed by anyone familiar with her history. Indeed, I’m not sure anyone watching Ammonite without prior knowledge of Anning would really think she was anyone especially important. By the end of the first act, Ammonite is basically done with palaeontology, and the rest of the narrative could easily be about any other downtrodden 19th century female professional you care to name or invent.
Second, yes, it’s time to address that topic: the decision to make Anning a character in a fictionalised romance rather than tell a component of her real history. Ammonite is only a Mary Anning film in the loosest sense: it has the right character names, the right location and the basic Anning backstory, but that's about it. Along with speculating about Anning’s sexuality (we have no data at all on Anning’s romantic interests), her character is also changed. We don’t know much about Anning’s personality, but historic notes - such as those cited in Deborah Cadbury’s 2000 book The Dinosaur Hunters - imply a very different character to that invented for the film. Quotes about later-life Anning describe a patient, kindly woman grateful for shop patrons, for example, which is a world apart from the icy, blunt Anning of Ammonite. The biggest historic casualty of the film, however, is not Anning, but her love interest, Charlotte Murchison. In real life, Murchison was a well-travelled, experienced geologist of great professional inspiration and importance to her geologist husband - the eventual Director-General of the British Geological Survey, Roderick Murchison. She was also a trailblazer for women’s rights in science, protesting against Charles Lyell for the right for women to attend scientific lectures. Ammonite’s Charlotte Murchison, in contrast, is a character defined only by her relationships with other people: a lonely wife grieving over the loss of her child, left with Anning in the hope that she may take up geology as a hobby. It’s certainly true that Ammonite is about an intelligent, important woman being brushed under the carpet of society, but there’s a meta quality to this that wasn’t expected.
It’s in this area that I find myself most conflicted about Ammonite. On the one hand, Ammonite’s narrative and the real stories of Anning and Murchison draw similar conclusions about sexism and classism, so the deviations from history might not matter too much: the imagined and real history meet, more or less, to impart the same message. On the other, the real Anning story is not only unique (and thus more interesting than any imagined drama) but also much more powerful. Women like Anning and Murchison literally changed the course of history and their lives couldn’t be more relevant to modern concerns: feminism, societal inequality, the power of rich white men, and the issue of privilege are all inescapable truths of their biographies. Ammonite replaces some of these with an equally important cause - LGBTQ representation - but - to our knowledge - this was never Anning’s fight, and it’s strange that she’s now been pushed as a figurehead in this movement too. As Riley Black mused in her excellent piece on Ammonite last year, we are at risk of asking Anning to carry too much, and thus diluting her real importance and legacy. The irony of Ammonite, a film that seemingly celebrates Anning, sidelining her scientific and intellectual achievements so it can impress its own narrative and significance onto her life is not lost on me. It’s not quite the same as writing her out of history like so many Victorian gentlemen scientists, but it’s certainly another example of people using her reputation for their own ends.
And that’s where Ammonite leaves me: a commendable historic drama with some great attention to characterisation and some great performances, levelled against disappointingly little interest in the richness of its source material and a confused, perhaps even self-defeating relationship with its principal characters. For all this, I still think Ammonite is, on balance, a Good Thing. While not delivering the sweeping Anning biopic many palaeontologists and historians might want to see, it still promotes - however contrarily in light of historic facts - the importance of women in science, gives a general idea of who Mary Anning was, and may spark some important thoughts and conversations among more interested audience members. And, lest we forget, behind all the talk about representation and legacies is a good film that can be watched and enjoyed, and it's OK to switch our brains off from time to time. Ultimately, whether you should check out Ammonite will depend on what you want from this picture. If you can get past the fact that Ammonite is little more than watered-down palaeontological fanfiction (perhaps the most niche genre in cinema), you may enjoy it. If you're unable to leave your scientific and historical brain at the door, this might not be for you. Whichever of these opposing views you land on, I agree with you.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
The giant elasmosaurid Albertonectes vanderveldei forages for invertebrates and small prey deep underwater. From this posterolateral view, the vertical tail fin is unmissable - but should I have drawn it as a horizontal fluke instead? Welcome to yet another challenge for restoring the life appearance of plesiosaurs!
It’s fair to assume that, when painting or sculpting plesiosaurs, most palaeoartists favour attention to their heads, necks and flippers, these being the most characteristic parts of their anatomy and the components most of our audience will focus on. Plesiosaurs are, of course, famously tricky animals to restore with any degree of certainty, their body plans being entirely unlike anything alive today, their mechanism of swimming - underwater flight with two sets of flippers - being the source of much debate and controversy, and their fossil record giving us few direct insights into their soft-tissue anatomy. So here’s more good news: in recent years another part of plesiosaur anatomy has joined their flippers and necks in artistic and scientific contention, their tails.
Plesiosaur tails have traditionally been little more than afterthoughts in palaeoartworks: cone-shaped structures tapering from the body without much in the way of interesting features. In the last decade or so, however, research interest in plesiosaur caudal anatomy and the resurrection of certain historic observations (e.g. Dames 1895; Smith 2007, 2013; Wilhem 2010; Sennikov 2015, 2019; Otero et al. 2018) has seen plesiosaurs with more interesting tails making regular palaeoartistic appearances. This artistic shift initially showed plesiosaur tails having fish- or ichthyosaur-like vertical fins but, more recently, newer research has made a case for another configuration, horizontal tail flukes (note the terminological distinctions), and such reconstructions are now also appearing with regularity. These fins and flukes, it must be stressed, are not mere whimsy or speculation but based on osteological correlates for some kind of extensive skin structure around the tail tip, and we should clarify that no-one thinks these represent a hitherto unappreciated propulsive organ - plesiosaurs do not suddenly have five ‘engines’ for swimming. Instead, these fins or flukes surely represent devices to act as some kind of rudder or stabilising aid, perhaps helping to offset the impact of swimming around with those giant necks and heads.
Recent skeletal reconstructions of plesiosaurs (or parts thereof) showing different interpretations of caudal rudder anatomy. It is increasingly common to see these features in plesiosaur skeletals and this compilation image could be much bigger, but I've restricted it to those from papers specifically making cases for fins or flukes.
One thing is clear: the evidence for tail rudders in plesiosaurs is pretty strong in several plesiosaur clades and any credible palaeoart of these animals should show them with some kind of fin, flipper or fluke at the tail tip. But, clearly, our current conflicting interpretations of plesiosaur tail anatomy can’t both be correct. I’ve gone back and forth on these ideas in my plesiosaur art over the last couple of years and decided that it was time I looked into this in more detail. What is the evidence for tail fins or flukes in plesiosaurs, and which is most compellingly argued for? Can we even make a call on this topic at the moment? Let's find out.
One of several plesiosaur artworks I've drawn recently showing a fluked tail, rather than (as was my previous preference) a fin. This is Plesiosaurus dolichodeirus scavenging a pterosaur carcass, because plesiosaurs are meant to eat pterosaurs in palaeoart, consarnit.
Something old is new... etc. etc.
First, let's briefly familiarise ourselves with the history of this controversy. It may seem that notions about plesiosaur caudal fins and flukes are part of the Brave New World of changing up old reconstructions of fossil reptiles for radical new ones, but that’s not the case here. Actually, osteological features and soft-tissue remains evidencing caudal rudders in plesiosaurs were identified in the 19th century, and even the dichotomy of interpretation between fins vs. flukes is over 100 years old. Our modern discussions of these concepts are a revival of relatively early investigations into plesiosaur functional morphology and life appearance.
The holotype of Seeleysaurus guilelmiimperatoris, famous for having a soft-tissue outline around its tail. This specimen was the only plesiosaur fossil on record with direct bearing on the fin vs. flipper controversy, but the tail soft-tissues have been painted over and are no longer accessible for study. Yeah, frustrating, right? From Dames (1895).
The concept of plesiosaur caudal fins was first broached by Richard Owen, who proposed that lateral compression of the terminal tail vertebrae of Archaeonectrus rostratus indicated a caudal fin (Owen 1865). Owen’s observation had little impact on reconstructions of plesiosaurs at the time, but seeming confirmation of his prediction arrived a few decades later when, in 1895, a specimen of the Jurassic plesiosaur Seeleysaurus guilelmiimperatoris was described with a soft-tissue outline around much of tail tip (Dames 1895). To date, this specimen remains the only fossil on record that provides direct confirmation of a caudal fin or fluke but, alas, it can no longer be investigated or even validated as the soft tissue component of the fossil has been painted over. This presents a problem deeper than merely obscuring the body outline. Many marine reptile specimens were often ‘improved’ with forged soft-tissues and realigned bones by 19th century preparators such that anything especially amazing and interesting - like the only known plesiosaur tail fin outline, for instance - really needs verification from modern researchers to be accepted as genuine. I’m not aware of any plesiosaur experts who consider the Seeleysaurus soft-tissues especially suspicious, but this inescapable caveat hangs around any discussion of this specimen: we can only put so much stock in any interpretation of it, modern or historic. This said, scholars of the late 1800s certainly regarded the body outline as genuine, leading to a few late 19th and 20th-century palaeoartworks in which Seeleysaurus and other plesiosaurs sported tall, diamond-shaped tail fins. Among the most famous of these were the reconstructions published in Wilhelm Dames' 1895 Die plesiosaurier der süddeutschen Liasformation and the generic pliosaurid reconstruction published by Newman and Tarlo (1967).
Neither finned nor fluked plesiosaurs really caught on in historic palaeoart, but finned reconstructions almost did. Here's a rarely seen example: Woodward's (1896) reconstruction of "Plesiosaurus" macrocepahlus, from A Guide to the fossil reptiles, amphibians, and fishes in the Department of Geology and Palaeontology of the British Museum (Natural History).
Not everyone accepted Seeleysaurus as evidence of caudal fins in plesiosaurs, however. Both Fraas (1910) and Wegner (1914) felt that their caudal skeletons were indicative of a horizontal fluke, citing the absence of a tail bend, the dorsoventral flexibility of the vertebrae, and the size of the caudal ribs as evidence of this feature. I’m not aware of any historic reconstructions showing this configuration (this is not to say that none exist, of course) and assume that fluked plesiosaurs gained even less traction in technical literature and artwork than their finned counterparts. This left most academic and artistic consideration of plesiosaur caudal anatomy assuming featureless, tapering tails for the next century, despite the continued cataloguing of peculiar caudal anatomy in plesiosaurs - most notably, their fused terminal tail vertebrae, recalling the pygostyles of birds (see review by Smith 2013). Today, such structures are considered part of the evidence package for a caudal rudder, but this significance seems to have been mostly overlooked in older publications.
Evidence for fins, evidence for flukes
The story of plesiosaur caudal rudders is thus one of three parts: an initial set of pro-rudder proposals and observations; a period of relative disinterest in the idea; and our modern era of belated engagement with those original hypotheses. This latter stage began about 10-15 years ago when observations made by Owen and Dames were resurrected and augmented by modern plesiosaur experts to make a case for plesiosaur tail fins. Perhaps the most in-depth investigations supporting finned tails to date are those by Benjamin C. Wilhelm (2010) and Adam Stuart Smith (2007, 2013), both of whom looked at the tails of Jurassic plesiosaurs and identified features also seen in swimming animals with vertical caudal fins. Using the relatively completely known tails of the cryptoclidids Cryptoclidus and Muraenosaurus, Wilhelm (2010) catalogued a suite of anatomies correlating with vertical fin lobes, including a relatively large neural spine close to the tail tip (the 17th caudal vertebra in Cryptoclidus); several neural spines with expanded ends; a shift in the orientation of the neural spines at the tip of the tail (from posteriorly-directed to anteriorly-directed) and lateral compression of the terminal caudal vertebrae (see diagram, above). These features recall the finned tails of mosasaurs, Triassic ichthyosaurs and thalattosuchians (e.g. Lindgren et al. 2013; Renesto et al. 2020), where enlarged neural spines mark the start of a caudal fin and shifting neural spine orientations characterise the vertebrae embedded in the fin itself. As with tail-finned reptiles and fish, plesiosaur tails had increased flexion in two regions, both at the tail base and immediately anterior to the enlarged and reorientated neural spines. A slight vertical peduncle - a thinning of the tail structure to minimise its drag profile when being moved through water - was also identified anterior to the enlarged neural spine region. By analogy with the preserved soft-tissues of other marine reptiles, Wilhelm’s study allowed for a prediction of the possible fin outline for cryptoclidids: a small triangular lobe with a concave posterior margin on the upper side of the tail (above). This is quite different from the diamond-shape interpreted by Dames and others from the Seeleysaurus fossil, but Wilhem’s thesis argues that a large ventral lobe is not suggested by that fossil and we can infer - if Wilhelm’s reconstruction is correct - that only a small amount of material is missing from the dorsal margin (see diagram, below).
Advocates for plesiosaur tail fins highlight similarities between their tail anatomy (see illustrations, above) and those of certain finned marine reptiles, such as early ichthyosaurs and mosasaurs. There are indeed obvious similarities, although the plesiosaur fin supports are clearly far less developed than the examples shown here - a reflection, almost certainly, of tails being adapted for locomotion vs. those adapted for ruddering. Images from Renesto et al. (2020) and Lindgren et al. (2013).
Although working with more fragmentary material, Smith (2007, 2013) found similar features to those reported in Benjamin Wilhelm’s thesis, as well as further evidence of finned tails. This included lateral compression at the tail tip of Rhomaeleosaurus, as well as peculiar ‘node’ vertebrae that might indicate a zone of flexion or even a slight downturn of the tail tip - another feature of finned tails. A wedge-shaped vertebra was noted in another rhomaleosaurid, Macroplata, which might indicate a slight tail downturn, although this taxon curiously lacks laterally compressed distal vertebrae. Agreeing with Benjamin Wilhelm’s thesis, as well as Wilhelm and O’Keefe’s (2010) examination of further caudal material, Adam's study concluded that the flexibility evidenced in the proximal tail region would allow the caudal fin of plesiosaurs to augment steering and stability.
A few years after these works reignited interest in plesiosaur caudal rudders, artistic and academic champions of fluked plesiosaurs also resurrected, retooled and expanded the initial observations and arguments made by Fraas and Wegner (e.g. Sennikov 2015, 2019; Otero et al. 2018). At the core of these proposals were ideas that plesiosaur tails were more convergent with those of whales and manatees than with swimming reptiles or fish, and thus suited to dorsoventral motion and sporting horizontally-aligned soft-tissues at their tips. Some researchers have endorsed this idea by including fluked reconstructions in their papers (e.g. Sachs et al. 2016).
Were plesiosaur tails reptilian variants of sirenian-like caudal anatomy? Again, there are similarities, especially in the great width of the vertebrae. In lieu of decent sirenian skeletons online, I've borrowed these Florida manatee (Trichechus manatus) skeleton replica images from Bone Clones.
Some of the evidence for plesiosaur flukes is the same as that used for finned reconstructions, such as the two zones of flexion in the plesiosaur caudal skeleton, and the recognition of a distinctive, often stiffened vertebral region at the end of the tail (Sennikov 2015, 2019). This is representative of fluked and finned tails having functional similarity, each essentially being the same thing operating in different planes. Fluke-specific evidence includes the presence of long caudal ribs along much of the tail length which, in making some vertebrae wider than tall, are suggested to create a relatively wide, flat tail with restricted lateral movement (Sennikov 2015, 2019; Otero et al. 2018). Otero et al. (2018) noted that the tips of the caudal ribs of Aristonectes are fibrous for potential attachment of extensive soft-tissue, potentially implying a much wider tail than shown by osteology alone (something seemingly confirmed by soft-tissue data of Mauriciosaurus). The absence of convincing downturned tail tips has also been flagged up, even for taxa with ‘node’ vertebrae (Otero et al. 2018), and has been negatively compared against the finned condition of ichthyosaurs and marine crocodylomorphs (Sennikov 2019). The small size or absence of chevrons, and the low, variable orientation of caudal neural spines, are features thought to have allowed the fluke to move vertically and independently of the rest of the tail (Otero et al. 2018; Sennikov 2019). The plesiosaur torso - a pachyostotic, relatively inflexible trunk - has also been regarded as similar to that of sirenians and thus potentially indicative of a fluked tail (Sennikov 2019). For the fluke model to be correct, of course, the traditional interpretation of Seeleysaurus having some kind of tail fin has to be wrong. Sennikov (2019) states that this is indeed the case, and provides an alternative view where the Seeleysaurus soft-tissues represent a horizontally-aligned structure within which the vertebrae have fallen over. The asymmetry of the fluke as preserved is explained as a result of vertebral displacement, soft-tissue decay and incomplete fossilisation.
Problematic as it is, proposals of plesiosaur rudder shape have to contend with the now-lost Seeleysaurus data to some extent. Some fin models posit that not much has happened to the caudal region of the Seeleysaurus holotype, whereas the fluke model requires a little more modification. While the fin model seems simplest, the fluke version hardly seems implausible.
Fins vs. flukes, head to head (er... tail to tail?)
Having briefly reviewed this evidence, can we see which of these models seems strongest? I stress my word choice in that sentence: our understanding of plesiosaur soft-tissues is not yet sufficient to make claims of absolute certainty, so any conversation on them should be peppered with appropriate caveats and considerations. This is a case where it’s much easier to be a researching scientist, where “we don’t yet know” is a perfectly acceptable response, than a palaeoartist, where you have to come down on one side of a debate or another. So, with the proviso that I’m not sure we can really make a definitive call yet… and with some urging of readers to check the papers mentioned above for themselves to make up their own minds... and with appropriate caution… and having now run out of ways to stall writing this sentence... I find myself... more persuaded by arguments for a tail fin at present. The fin hypothesis seems to explain more of the peculiar anatomy of plesiosaur tail tips than its rival, and it presents an overall simpler, and thus more likely correct, interpretation of our available data. The highlighted similarities between plesiosaur tails and those of certain other fin-tailed swimming reptiles are prudent observations not yet accounted for in the fluke model and, when thinking about the evolution of such features, it seems a shorter developmental distance for a plesiosaur to evolve a fin than a fluke. I’m especially thinking of the switch from primarily lateral to vertical spinal flexion, the former being ancestral to reptiles and thus common to many in swimming reptile species. Significant vertical caudal flexion has developed among diapsids of course, in birds, and some swimming birds even use their tail fans in a rudder-like fashion (Felice and O’Connor 2014), so we shouldn’t rule out this capability for plesiosaurs entirely. Moreover, this is not to say that the observations from Team Fluke are redundant. Maybe the wide, potentially muscular tails of plesiosaurs were indeed capable of an unusual amount of vertical motion, even with a fin? If, indeed, plesiosaur tails operated as ruddering aids, some surely had a big job on their hands: there’s no way moving those giant necks and heads didn’t have a tremendous rotational impact when swimming and their tails may have had an important role in keeping their owners on course. A weedy tail with limited mobility in either plane might not have been up to that task. It would be neat to see simulations of this sort of thing.
I also agree with Wilhelm’s (2010) assessment of the Seeleysaurus tail outline, which looks - so far as can be seen in Dames’ (1895) illustrations, more like a vertical structure than a horizontal one. I’m not sure how much stock we should place in interpretations of the drawings of this specimen given that the original soft tissues are now inaccessible, but, for what it’s worth, the relatively neat margins and asymmetry of the tissue outline, the general position of the vertebrae and location of possible ‘fin’ tissues above the tallest neural spines look consistent with a small dorsal lobe (sensu Wilhelm 2010), while a fluked interpretation relies on a fair amount of distortion from decay and disarticulation. But there is certainly plenty of ambiguity around that specimen and I could be convinced otherwise. Needless to say, more plesiosaur specimens with soft-tissues preserved around the tail tip would be incredibly useful in this discussion.
Plesiosauria was a diverse group with a huge array of body proportions, sizes and lifestyles. Did the gigantic pliosaur Kronosaurus queenslandicus share the same caudal rudder shape and size as Cryptoclidus? We need more research to say either way.
Of course, this tentative endorsement of vertical fins comes with many caveats. Despite the antiquity of proposals that plesiosaurs bore a soft-tissue rudder on their tails, these discussions are still in their infancy. Very few studies have directly addressed this topic and our analyses have, thus far, been largely limited to qualitative descriptions and comparisons with other vertebrates. Our taxon sampling is also relatively limited and has not yet incorporated the most extreme examples of plesiosaur size or body plans. I can’t be the only one wondering what the extremely long-necked elasmosaurids and giant pliosaurids were doing with their tails and, given the innumerable studies linking tail shape to ecology in swimming animals, it’s not crazy to assume plesiosaurs of different lifestyles might have had differently shaped rudders. We may even already be finding evidence of such variation, it being observed (for example) that elasmosaurids may not have had the mobile terminal tail region identified in some cryptoclidids (Otero et al. 2018). Moreover, might plesiosaurs have done something unusual with their tails, such that the fin vs. fluke dichotomy is an oversimplification of a more complex organ befitting their unique swimming style? Could other structures - keels or projecting stabilisers - have augmented a larger fin? It will be interesting to see what further research reveals and, in particular, what quantified comparisons between the caudals of plesiosaurs and those of fluked and flippered animals have to tell us. Many of the features discussed above are relatively subtle - a taller neural spine here, a slight wedging of the centrum there - and careful, quantified comparisons of a suite of features with modern and extinct analogues may help us distinguish whether fins or flukes are the better tail rudder model, or if such analogues are useful guides at all.
And that, with one foot off the fence on this matter, but still not feeling like a conclusion has really been reached, is where we'll leave off for now. Hopefully, more definitive evidence for plesiosaur tail morphology will appear soon but, until then, the best thing artists can do is not take the comments above as gospel: check out the papers discussed here (most are open access or otherwise available online) to make up your own mind which is the better-supported model. I suspect I'm still going to remain uncertain about my plesiosaur tails for some time, but there's comfort knowing that this isn't really something we can be especially confident about until more data is gathered.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Dames, W. (1895). Die plesiosaurier der Süddeutschen Liasformation. Abhandlungen der
Königlich Preussischen Akademie der Wissenschaften zu Berlin 1895, 1–81.
Felice, R. N., & O’connor, P. M. (2014). Ecology and caudal skeletal morphology in birds: the convergent evolution of pygostyle shape in underwater foraging taxa. PLoS One, 9(2), e89737.
Fraas, E. (1910). Plesiosaurier aus dem oberen Lias von Holzmaden, Palaeontographica, 57, 3–4, 105–140.
Lindgren, J., Kaddumi, H. F., & Polcyn, M. J. (2013). Soft tissue preservation in a fossil marine lizard with a bilobed tail fin. Nature Communications, 4(1), 1-8.
Newman, B. & Tarlo, B. (1967). A giant marine reptile from Bedfordshire. Animals, 10, 61-63
Otero, R. A., Soto-Acuña, S., & O'keefe, F. R. (2018). Osteology of Aristonectes quiriquinensis (Elasmosauridae, Aristonectinae) from the upper Maastrichtian of central Chile. Journal of Vertebrate Paleontology, 38(1), e1408638.
Owen, R. (1865). A monograph on the fossil Reptilia of the Liassic Formations. Part 3. Sauropterygia. Monograph of the Palaeontographical Society, 17, 1–40, pl. 1–16.
Renesto, S., Dal Sasso, C., Fogliazza, F., & Ragni, C. (2020). New findings reveal that the Middle Triassic ichthyosaur Mixosaurus cornalianus is the oldest amniote with a dorsal fin. Acta Palaeontologica Polonica, 65(3), 511-522.
Sachs, S., Hornung, J. J., & Kear, B. P. (2016). Reappraisal of Europe’s most complete Early Cretaceous plesiosaurian: Brancasaurus brancai Wegner, 1914 from the “Wealden facies” of Germany. PeerJ, 4, e2813.
Sennikov, A. G. (2015). New data on the herpetofauna of the Early Triassic Donskaya Luka locality, Volgograd Region. Paleontological Journal, 49(11), 1161-1173.
Sennikov, A. G. (2019). Peculiarities of the Structure and Locomotor Function of the Tail in Sauropterygia. Biology Bulletin, 46(7), 751-762.
Smith, A. S. 2007. Anatomy and systematics of the Rhomaleosauridae (Sauropterygia: Plesiosauria). Unpublished PhD thesis, School of Biology and Environmental Science, National University of Ireland, University College Dublin.
Smith, A. S. (2013). Morphology of the caudal vertebrae in Rhomaleosaurus zetlandicus and a review of the evidence for a tail fin in Plesiosauria. Paludicola, 9(3), 144-158.
Wegner, T. (1914). Brancasaurus brancai Wegner, ein elasmosauride aus dem Wealden Westfalens. Borntraeger.
Wilhelm, B. C. (2010). Novel anatomy of cryptoclidid plesiosaurs with comments on axial locomotion (Doctoral dissertation, Marshall University Libraries).
Wilhelm, B. C., & O'keefe, F. R. (2010). A new partial skeleton of a cryptocleidoid plesiosaur from the Upper Jurassic Sundance Formation of Wyoming. Journal of Vertebrate Paleontology, 30(6), 1736-1742.
When it comes to imagining aggressive behaviour between large Mesozoic theropod dinosaurs, the main game in town is head-biting: individuals grappling with one another by locking jaws around each other’s faces (Tanke and Currie 1998; Peterson et al. 2009). Such actions are well-evidenced by the sometimes significant trauma found on theropod skull fossils, including tooth marks, gouges, and even teeth embedded in facial bones. Clearly, head-biting was a dangerous, bloody activity that could lead to significant injury, and we have to wonder if theropods might have had means of physically settling their differences without tearing each other’s faces apart. After all, while some modern animals don’t have much behavioural grey space between ‘idle’ and ‘psycho’, many species employ means of fighting that prevent, or at least diminish, the need for more violent encounters.
One non-lethal means of theropod interaction explored in dinosaur literature is head-butting, where the various crests, horns and bosses adorning their skulls are transformed into bludgeons, clubs and shoving aids. We don’t see or read much about theropods employing these behaviours in dinosaur popular culture despite its obvious appeal - if you don't find the idea of heckin' big predatory dinosaurs shoving each other around with their heads at least a little bit awesome, you might want to see a doctor. But, actually, theropod head-butting has a reasonable foothold in dinosaur technical literature, with some studies even dedicated to determining its feasibility.
Among the earliest proponents of theropod head-butting were Robert Bakker and Gregory S. Paul, who've been illustrating this behaviour and the anatomy related to it for decades. From left to right: Bakker's (1986) diagram of theropod head ornaments related to head-butting; Paul's (1987, but published in 1988) fantastic flank-butting Ceratosaurus; and a more recent Bakker work showing head-butting Tyrannosaurus from the Beyond Bonesblog.
So far as I’m aware, the concept of theropod head-butting was a product of the Dinosaur Renaissance, perhaps an unsurprising occurrence given that it involves regarding theropods as relatively sophisticated, behaviourally-complex animals rather than the pea-brained dullards of pre-Renaissance times. Two of the most influential figures of that revolution - Bob Bakker and Gregory S. Paul - were strong advocates for theropod head-butting. Their famous books The Dinosaur Heresies (Bakker 1986) and Predatory Dinosaurs of the World (Paul 1988) both proposed that theropods engaged in these behaviours (above). Bakker’s take interpreted the stout ridges and crests of theropods such as Allosaurus and Tyrannosaurus as weapons for butting bouts and stressed their significance as nonlethal fighting devices. Paul considered head-butting likely for most theropods with robust cranial ornaments, from the elaborately horned Ceratosaurus and Carnotaurus to the massively-skulled tyrannosaurs. Paul favourably compared the various horns and bosses of such animals to those of giraffes (which, of course, are renowned head clubbers) and has maintained such views in more recent works (e.g. Paul 2016). I’m curious as to whether Bakker and Paul's affinity for drawing theropods played into the formation of these ideas, as they both were among the strongest critics of pre-Renaissance artists for not capturing the diverse morphologies and ornaments of theropod skulls and produced many artworks showing more accurate theropod depictions. Might their more precise renderings of theropod skulls have made them ponder the function of the bosses, ridges and crests they were illustrating?
In subsequent decades theropod head-butting has been mentioned semi-often by theropod workers. A minority of workers have been negative and dismissive of the idea (e.g. Rowe 1989; Molnar and Farlow 1990), but it’s mostly brought up in a confirmatory sense. Most discussions have centred around a few clades - tyrannosaurids (e.g. Bakker 1986, Paul 1988, 2016), carcharodontosaurids (e.g. Sereno and Brusatte 2008; Cau et al. 2013) and especially abelisaurids (e.g. Novas 1989; Mazzetta et al. 1998, 2009; Hieronymus 2009; Snively et al. 2011; Delcourt 2018; Cerroni et al. 2021) - and we now have a suite of typical anatomical features identified as potential head-butting structures, as well as dedicated biomechanical investigations into the plausibility of theropod skull combat (Mazzetta et al. 1998, 2009; Snively et al. 2011; Xing et al. 2015). Evidently, the concept of theropod head-butting is not - as is sometimes the case - an over-eccentric hangover from a particularly adventurous period of dinosaur research, but a legitimate and current behavioural hypothesis supported by a growing amount of data.
The postorbital bone of the carcharodontosaurid Eocarcharia dinops and its prominent orbital boss: a structure made for lateral head-butting? From Sereno and Brusatte (2008).
Let's get into this a little more: what, exactly, are the structures theropod workers are linking to head-butting? As will be well-known to anyone reading this blog, theropods were prone to augmenting their skulls with ridges, bosses, crests and horns, most commonly along the top of the snout and above their eyes. It’s among these that we find potential head-butting structures, but it's not thought that any and all ornaments were suitable for head-butting. Some theropod crests were surely too delicate for physical aggression, probably even allowing for the strengthening properties of their overlying skin tissues (see below). It’s hard to believe that the thin, tall crests of Dilophosaurus were being smacked into other animals, for example, and they instead surely served a purely signalling role (Bakker 1986; Rowe 1989). Indeed, studies of the Dilophosaurus-like crests of Sinosaurus found that they were prone to structural failure under mechanical loading (Xing et al. 2015), and likely ill-suited to weaponisation.
The 'Trix' specimen of Tyrannosaurus, on display in Glasgow 2019, showing the massive postorbital bosses which dominate the posterior skull region of this species. Researchers have associated these with head-butting behaviour, which seems reasonable given their structure, location, and the general grumpy attitude evidenced for Tyrannosaurus.
Head-butting structures are better inferred from the sizeable, prominent and robust components adorning certain theropod skulls. In carcharodontosaurids, ornament indicative of head-butting consists of swollen, laterally-prominent bosses above their eyes (Sereno and Brusatte 2008) or, more unusually, dorsally-prominent domes in the same region (Cau et al. 2017). Unfortunately, taxa with these features - Eocharchia and Sauroniops - are not especially well-known so we can’t see if these features were somehow echoed across the rest of the skull: might they have sported other enhanced bosses and ridges as well? This is not a problem for tyrannosaurids, of course, which have skulls of extreme familiarity to theropod researchers. Both Bakker (1986) and Paul (1988, 2016) have linked the robust nasal bones (forming the top of the snout), lacrimal cornual processes (horns in front of the eye) and postorbital bosses (swollen, rounded structures behind the eye) of tyrannosaurs with head-butting behaviour. Predictions of armoured skin around their eyes and snout (Carr et al. 2017) are consistent with this idea, too. Tyrannosaurus needs special mention here for its uniqueness among theropods - even other tyrannosaurs - in having flattened its lacrimal cornual processes into swollen, highly-textured expansions of the skull roof and inflated its postorbital bosses to a great extent, sometimes even capping the latter with osteoderms (Carr et al. 2017). The enlarged postorbital bosses are among the tallest parts of the skull and also project outward from the eye socket by a considerable margin. Significant rugosities and other properties of these lacrimal and postorbital features indicate a covering of especially thick, toughened skin, but no true horns (despite what you see in lots of artwork and films, there’s nothing pointed or horn-shaped about the ornament of big T. rex specimens). If Bakker and Paul are correct in Tyrannosaurus having a head adapted for head-butting, it’s difficult not to see this reconfiguration as the skull of a theropodan bulldozer: a flattened, shovel-shaped cranium to heave and bash other animals around with.
Tyrannosaurus is unusual among tyrannosaurids for lacking a hornlet in front of the eye, but it compensated for this with armoured cranial skin and two prominent postorbital bosses. Bolted to something like six or more tonnes of angry tyrannosaur, this must have made for one heck of a battering ram. So let's see... if there's BRONTOSMASH!, ArSUMOitherium, then this must be... TYRANNOBUTT? Yes, make it so.
Neither tyrannosaurs nor carcharodontosaurid head-butting has received much in the way of dedicated research attention, however, leaving our best insights into this behaviour coming from abelisaurs, especially Carnotaurus. The head-butting potential of these short- and gnarly-faced, sometimes horned theropods has been remarked on for decades (e.g. Paul 1988; Novas 1989; Mazzetta et al. 1998) and, via Carnotaurus, encouraged several rounds of dedicated biomechanical investigation (Mazzetta et al. 1998, 2009; Snively et al. 2011; Méndez 2012). These initially regarded Carnotaurus cranial combat as a brutal affair, with spinal dampening allowing combatants to impact their heads at rhinoceros-grade speeds of 20 kph (Mazetta et al. 1998). But subsequent investigations have curbed these ideas somewhat, finding that Carnotaurus skulls were incapable of sustaining strong forces during simulated head-butting (Mazetta et al. 2009). Instead, their skulls were better suited to low-velocity impacts, blows that targeted softer parts of animal bodies (e.g. flank butting), or maybe even quasi-static bouts of head-shoving (Mazzetta et al. 2009). This isn’t too surprising: Carnotaurus is a relatively robustly-skulled theropod, but its cranium is still a relatively vacuous, air-filled structure adaptively balancing maximised strength and minimised weight. Compared to the massive and thickened skulls of more dedicated headbutters, theropod skulls look relatively delicate. Abelisaur necks are no longer considered shock absorbers for running impacts either, but their recognition as strong, powerful elements means they still have a role in head-butting hypotheses (Méndez 2012): the utility of a powerful neck in energetic, sustained use of a weaponised head is obvious (Delcourt 2018).
Epidermal correlates covering the heads of abelisaur species, from Delcourt (2018). Carnotaurus stands out with its indications of especially reinforced, cornified snout and horn tissues - a 'carnivorous bull' indeed.
Further information on abelisaur head-butting stems from their facial skin. The rugose surface textures of abelisaur skulls are indicative of particular skin types, such as cornified sheaths (think bird beaks and cow horns), cornified pads (muskox heads) or thickened, armoured dermis (rhino and hippo skin) (Sampson and Witmer 2007; Hieronymus 2009; Delcourt 2018; Cerroni et al. 2021; Hendrickx and Bell 2021). These are exactly the tough, reinforced skin types we’d expect to see in animals that used their heads as clubs or shoving implements, and it’s been noted that these dermal types, in extant animals, correlate with intraspecific fighting (Hieronymus 2009). We’ve touched on these data before, discussing that abelisaur faces may have looked quite different to their skulls thanks to thick skin. This is especially so for Carnotaurus, which had a snout covered by an extensive cornified pad. Because the evidence for thick, armoured skin on abelisaur faces is especially prominent compared to other theropods, it seems reasonable to infer a behavioural emphasis on head-butting within the group.
Carnotaurus sastrei, a species that - among theropods at least - is probably the most head-butting adapted of all. Exactly how its head, equipped with a cornified pad and sheathed horns, was used in aggressive behaviours has been interpreted differently over the years and boils down to assumptions of shock-absorption capability: was it a high-impact or low-impact headbutter, a shover, or a flank-butter? Whichever it was, it certainly looked awesome.
But these neat dermal insights bring complications along with insight, adding important caveats to the biomechanical studies outlined above. We know from living animals that head-butting capability is best modelled by factoring both bony and soft-tissue data (e.g. Drake et al. 2016), and that such simulations can turn bone-only models of skull strength on their heads. So far as I know, no studies of abelisaur cranial strength in head-butting simulations have modelled their armoured and cornified skin tissues yet, and so we await such analyses before real confidence can be drawn about abelisaur head-butting capability. The same is true, of course, of any other theropod we’d want to investigate head-butting behaviour for. I'm sure I'm not the only one thinking such studies, and other insights into this relatively unexplored but fascinating facet of theropod palaeobiology, would be very welcome additions to dinosaur research.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Bakker, R. T. (1986). The dinosaur heresies: new theories unlocking the mystery of the dinosaurs and their extinction. William Morrow.
Cau, A., Dalla Vecchia, F. M., & Fabbri, M. (2013). A thick-skulled theropod (Dinosauria, Saurischia) from the Upper Cretaceous of Morocco with implications for carcharodontosaurid cranial evolution. Cretaceous Research, 40, 251-260.
Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7(1), 1-11.
Cerroni, M. A., Canale, J. I., & Novas, F. E. (2021). The skull of Carnotaurus sastrei Bonaparte 1985 revisited: insights from craniofacial bones, palate and lower jaw. Historical Biology, 33(10), 2444-2485.
Delcourt, R. (2018). Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific reports, 8(1), 1-12.
Drake, A., Donahue, T. L. H., Stansloski, M., Fox, K., Wheatley, B. B., & Donahue, S. W. (2016). Horn and horn core trabecular bone of bighorn sheep rams absorbs impact energy and reduces brain cavity accelerations during high impact ramming of the skull. Acta biomaterialia, 44, 41-50.
Hieronymus, T. L. (2009). Osteological Correlates of Cephalic Skin Structures in Amniota: Documenting the Evolution of Display and Feeding Structures with Fossil Data (Doctoral dissertation, Ohio University).
Mazzetta, G. V., Fariña, R. A., & Vizcaíno, S. F. (1998). On the palaeobiology of the South American horned theropod Carnotaurus sastrei Bonaparte. Gaia, 15(185), 192.
Mazzetta, G. V., Cisilino, A. P., Blanco, R. E., & Calvo, N. (2009). Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei. Journal of Vertebrate Paleontology, 29(3), 822-830.
Méndez, A. H. (2012). The cervical vertebrae of the Late Cretaceous abelisaurid dinosaur Carnotaurus sastrei. Acta Palaeontologica Polonica, 59(3), 569-579.
Molnar, R. E., & Farlow, J. O. (1990). Carnosaur paleobiology. In: Weishampel, D. B., Dodson, P., & Osmólska, H. (eds.). The Dinosauria. University of California Press. 210-224 pp.
Novas, F. E. (1989). Los dinosaurios carnívoros de la Argentina. PhD thesis, La Plata: Universidad Nacional de La Plata.
Paul, G. S. (1988). Predatory dinosaurs of the world: a complete illustrated guide. Simon & Schuster.
Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press.
Peterson, J. E., Henderson, M. D., Scherer, R. P., & Vittore, C. P. (2009). Face biting on a juvenile tyrannosaurid and behavioral implications. Palaios, 24(11), 780-784.
Rowe, T. (1989). The early history of theropods. Short Courses in Paleontology, 2, 100-112.
Sampson, S. D., & Witmer, L. M. (2007). Craniofacial anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology, 27(S2), 32-104.
Sereno, P. C., & Brusatte, S. L. (2008). Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger. Acta Palaeontologica Polonica, 53(1), 15-46.
Snively, E., Cotton, J. R., Witmer, L., Ridgely, R., & Theodor, J. (2011). Finite Element Comparison of Cranial Sinus Function in the Dinosaur Majungasaurus and Head-Clubbing Giraffes. In Summer Bioengineering Conference (Vol. 54587, pp. 1075-1076). American Society of Mechanical Engineers.
Tanke, D. H., & Currie, P. J. (1998). Head-biting behavior in theropod dinosaurs: paleopathological evidence. GAIA: revista de geociências, (15), 167.
Xing, L., Wang, Y., Snively, E., Zhang, J., Dong, Z., Burns, M. E., & Currie, P. J. (2015). Model-based identification of mechanical characteristics of Sinosaurus (Theropoda) crests. Acta Geologica Sinica, 89(1), 1-11.
It's a well-verified fact that the best dinosaurs to draw are the sauropods, exemplified here by the mighty Giraffatitan brancai. But the route to this modern realisation was a difficult one. When sauropods were first discovered no one wanted to restore them at all: their fossils were always in the hands of the wrong people, at the wrong time, and our first attempt at restoring their life appearance was many decades after their discovery. A strange story awaits.
Is there any shape more emblematic of prehistoric life than the silhouette of a sauropod? The combination of long neck, long tail, round body and four robust limbs is universal visual shorthand for anything to do with dinosaurs and, more broadly, extinct life, such that sauropods are bone fide superstars of palaeontological pop-culture. The world-renowned Sinclair logo, Gertie, the leads of The Land Before Time and The Good Dinosaur, the scene welcoming us to Jurassic Park… all sauropods.
Much of the most famous sauropod iconography is, as with so much palaeontological media, related to fully-fleshed sauropod reconstructions rather than their fossil bones, so we can consider much of their popular appeal lying within paleoart. Given the undeniable spectacle and wonder associated with sauropod fossils, where even single bones can be jaw-dropping museum centrepieces, we might imagine that early scientists and palaeoartists jumped at the chance to put flesh on the skeletons of these long-necked reptiles as soon fossil material revealed their basic shape. And yet, this was not the case. The story of how sauropods entered the palaeoart canon and became ambassadors for all things dinosaurian and extinct is a peculiar one, where a full half-century passed between their discovery and their palaeoart canonisation. This is a story of how combative attitudes and egos, missed opportunities and perceived failures in early palaeoartworks stymied life reconstructions of sauropods for much of the 19th century.
(The following post owes much to the history of sauropod research outlined by Mike Taylor (2010) and Mark Hallett and Matt Wedel (2016). Be sure to check these out if you want a more detailed perspective on 19th century sauropod science.)
Fragmentary beginnings
Our tale begins, of course, with the discovery and description of the first sauropods Cetiosaurus and "Cardiodon" by Richard Owen in 1841. As with all the first dinosaur discoveries, our first sauropod fossils were isolated bones from southern Britain. Little could be said about the life appearance of the animals they represented other than that they were enormous reptiles. On account of their size, it was assumed sauropods must have been aquatic and – like all giant sea creatures – carnivorous. Perhaps, it was wondered, they were the arch predators of Mesozoic oceans: the devourers of plesiosaurs and ichthyosaurs. Owen speculated that such a creature must have borne a well-developed caudal fin perhaps similar to that of an ichthyosaur, but this awesome vision of sauropods did not lead to any life reconstructions despite the cavalier reputation of early 19th century Europeans for restoring extinct animals (if you’ve been reading my blog since its origins, you may remember an article on this very topic).
We might wonder why this was the case. After all, animals like Megalosaurus were initially represented by little more material than Cetiosaurus, but they were still restored within just a few years of their discovery. The absence of early 19th century sauropod reconstructions shows, to my mind, some level of nuance about early Victorian palaeoartists. Though often characterised as wild and speculative, the early Victorian palaeoart canon was surprisingly conservative, mainly showing similar scenes populated by the same species in the same anatomical guises. Many of their depictions were also of animals with decent fossil representation like marine reptiles, pterosaurs and fossil mammals, such that the restoration of poorly-represented species, like dinosaurs, can be viewed as exceptional. Moreover, and in further defence of the first palaeoartists, dinosaur anatomy wasn’t a total unknown in the early 19th century. Several species had been attributed characteristic anatomical features such as nose horns, body armour, or distinctive teeth, so there was something to hang restorations around. But sauropods lacked even a basic defining feature, and thus had no way of being distinguished or characterised from other restored giant reptiles. Should vindictive time-travellers ever force early Victorians to draw fleshed-out sauropods, the result would probably be nothing more than generic, giant carnivorous reptiles, perhaps little different to some of John Martin’s whale-sized, lizard-like dinosaurs.
John Martin's frontispiece to G. F. Richardson's 1842 book Geology for Beginners, entitled Age of Reptiles. A lot of early 19th century dinosaur palaeoart was not especially attentive to details of anatomy and form, such that sauropods, if they had been illustrated, probably would have looked much like the (?)Megalosaurus in this piece.
The whale lizard flounders in the palaeoart doldrums
A major step towards understanding sauropod life appearance was made in the late 1860s when a large haul of Cetiosaurus bones were found in Oxfordshire, UK. Described by geologist John Phillips in the 1871 book Geology of Oxford and the Valley of the Thames, this partial skeleton included limb bones, ribs, limb girdles and many vertebrae, and thus provided our first decent insight into the sauropod body plan. In what seems like a cruel twist of fate, no neck or skull bones were recovered, denying knowledge of the most defining characteristic of the group for another few years. Nevertheless, Phillips had enough anatomy to start making the first relatively informed insights into sauropod appearance and behaviour, and he devoted several pages of his book to discussing the size, habits and even skin of Cetiosaurus. Much of what we now think about sauropods was prophesied here, with Phillips describing very large, scaly reptiles that lived on land and ate plants. He made a number of favourable comparisons to Iguanodon in his discussion of Cetiosaurus habits such that he may have imagined it as an especially gigantic large-bodied, quadrupedal herbivore in the vein of Waterhouse Hawkins’ Crystal Palace Iguanodon and Hylaeosaurus. It is curious, therefore, in light of this evident interest, that Phillips did not attempt some sort of reconstruction. He had enough bones to at least construct a decent skeletal diagram. But, OK - maybe reconstructions weren’t Phillip’s thing. With all this anatomical information, surely someone else picked up the baton?
John Phillips' quarry map of a partial Cetiosaurus skeleton found in 1869-70. A lot of the vertebrae and smaller bones are amalgamated into packages in this drawing, but the glut of material is obvious. This was the first significant insight into sauropod life appearance... but not much came of it among Victorian palaeoart. From Phillips (1871).
And it’s at this point that our story becomes rather strange. Cetiosaurus, which was by now as well represented as other animals routinely featured in palaeoart, continued to be snubbed by artists. We have to take a step back from sauropod palaeoart history and expand our scope to the discipline as a whole to understand why. Along with Cetiosaurus, a number of genuinely important, game-changing dinosaur discoveries had been made across Europe in the mid-1800s that included Scelidosaurus (the first complete dinosaur skeleton), Compsognathus, Hypsilophodon, and Archaeopteryx. All were represented by excellent fossils that dramatically enhanced our understanding of dinosaurs and had major implications for reconstructing their life appearance, and yet none were canonised into palaeoart of the day. If you look hard enough you may find some simple line drawings of Archaeopteryx here and there, but there were no lavish paintings, no sculptures or elaborate lithographs celebrating these new animals.
Evidently, Cetiosaurus itself wasn't being snubbed. These superior sauropod remains had arrived while European palaeoart was in a funk that would last several decades, an era when most new palaeoartworks featured restorations recycled from the early 1800s instead of novel reconstructions of newly-discovered species. A contributing factor to suppressed creativity in late 19th century European palaeoart was, ironically, the Crystal Palace Dinosaurs (Secord 2004; Nieuwland 2019). Although popular with the public, most academic response to the 1854 unveiling of these models was negative. Criticisms were many, focusing on their speculated elements, the juxtaposition of modern landscapes with extinct animals, and accusations that they were simply scaled up, monsterised living species. Several Crystal Palace reconstructions were also rapid embarrassed by new fossil data, such that these flagships of Victorian palaeoart were now misleading or confusing the public more than educating them. The Crystal Palace Dinosaurs had given scholars of the 1870s good reason to be sceptical of palaeoart, and the creation of new life reconstructions fell out of fashion for a generation.
American palaeoart to the rescue… sort of
The awakening of American palaeoartistry is one of the few major events that occurred in palaeoart history in the early-late 19th century. While American palaeoartists slowly found their feet, the discovery of excellent sauropod material in western states in the 1870s finally revealed the full spectacle of these amazing, unique animals. Among the first relatively well-known American sauropods was Camarasaurus, found in 1877 and described by Edward Drinker Cope in the same year. These specimens, at last, revealed something of the iconic sauropod neck. Scientists were finally impressed enough to reconstruct a sauropod skeleton, and the result was John A. Ryder’s composite mounted skeleton of a ribless, sail-tailed Camarasaurus, complete with a speculated skull. This mount was around 50 ft long and exhibited at a meeting of The American Philosophical Society in Philadelphia in December 1877, but no images of it were not published until 1914. Nor, for that matter, was it accompanied or followed by a life reconstruction. In fact, I don’t know that anyone has attempted to restore Ryder’s Camarasaurus, so I set aside an hour or so in preparation of this article to finally correct this important injustice. I'd love to see more from different artists - #justiceforRydersaurus!
OK, yes, John A. Ryder's 1877 Camarasaurus skeletal is a little bizarre, but the essence of sauropod form is there. It's a stone-cold crime that no one reconstructed the life appearance of this skeleton, so I've had a go here. Some attempt has been made to give the restoration a classical 19th century, Hawkinsian palaeoart flavour.
The absence of an 1877 Camarasaurus life restoration is all the more curious because of Cope’s association with this specimen. Cope is most famous for his feud with Othniel Marsh and the ‘Bone Wars’ period of American vertebrate palaeontology, but he was also one of the main instigators of American palaeoartistry. During the 1860s Cope produced iconic artworks of dinosaurs known from New Jersey which can be considered among the first flesh reconstructions of recognisable, basically anatomically accurate dinosaurs, as well as well-known depictions of taxa from the Western Interior Seaway. He was an advisor in the 1868 reconstruction of Hadrosaurus by Joseph Leidy and Benjamin Waterhouse Hawkins, and thus had connections to the first grandmaster of palaeoart himself, who was often based in the US for palaeoartistic purposes in the 1860s and 1870s. And yet, around all this, Cope never produced a published sauropod life reconstruction, and nor did his relationship with Hawkins yield any (published) sauropod illustrations. We may ascribe the latter to Hawkins’ dislike of Cope, whom he regarded as an overbearing, fussy collaborator. On at least one occasion he threatened to abandon a project entirely if Cope was involved (Bramwell and Peck 2008). Who knows how the early history of American palaeoart would have played out had these two giants of their disciplines been on better terms.
Othniel Marsh's 1883 restoration of Brontosaurus - a significant advance over the Ryder sauropod skeletal of just a few years prior.
Cope was, of course, not the only major player in the discovery of American sauropods. Indeed, Cope’s rival, Marsh, probably has a greater legacy with these animals, not only naming more species but also coining the name ‘Sauropoda’. Marsh named and described many now-iconic sauropods including Brontosaurus, Apatosaurus and Diplodocus, and in 1883 he published a Brontosaurus skeletal reconstruction that was, anatomically speaking, vastly superior to Ryder's Camarasaurus. Marsh was thus another person well-placed to facilitate the first artistic resurrection of sauropod life appearance… if only it weren't for his career-long disdain for palaeoart. Here's what he said on this topic in 1875:
I do not believe it possible at present to make restorations of any of the more important extinct animals of this country that will be of real value to science, or the public. In the few cases where materials exist for a restoration of the skeleton alone, these materials have not yet been worked out with sufficient care to make such a restoration perfectly satisfactory, and to go beyond this would in my judgment almost certainly end in serious mistakes. Where the skeleton, etc., is only partly known, the danger of error is of course much greater, and I would think it is very unwise to attempt restoration, as error in a case of this kind is very difficult to eradicate from the public mind… A few years hence we shall certainly have the material for some good restorations of our wonderful extinct animals, but the time is not yet.
(Marsh 1875, quoted in Dodson 1996, p.74)
Marsh’s criticism of palaeoart has a lot of implications for the development of the discipline in general, and almost certainly contributed to the delayed canonisation of sauropods. He held many of the best cards when it came to understanding sauropod life appearance but was the last person who would include a life restoration in a publication. Moreover, his reputation, influence, and longstanding criticism of palaeoart may have further dampened drives to restore newly discovered taxa in Europe, where some of his fiercest denouncement of palaeoartworks were expressed. Marsh’s views were surely a major contributor to the strange fact that the Bone Wars era - one of the most intense periods of discovery and analysis in early dinosaur history – was entirely bereft of associated palaeoart. And while Cope could, in theory, have picked up these palaeoartistic pieces, the ferocity of his feud with Marsh surely meant that he avoided reconstructing Marsh taxa, no matter how spectacular they were. There’s some irony in Marsh and Cope being so instrumental to our early conceptualisation of sauropods but that their various hang-ups - with each other, with other people, and with palaeoart - only kicked the can further down the road.
Finally, a life reconstruction! ...that everyone ignored
By this time - the late 1870s or early 1880s - sauropods had been known to science for about 40 years, with decent, restorable remains on record for at least half that time. Their gigantic size and spectacular anatomy were well appreciated and, thanks to Marsh, their skeletal anatomy had been committed to scientific literature. In these circumstances, surely someone was going to crack and attempt a flesh restoration? Yes, finally, someone did - but not in either of the historic homes of sauropods, Britain and the US. Rather, it was the French author Nicolas Camille Flammarion who provided the first (to my knowledge) sauropod life restoration in his 1886 book Le Monde Avant la Création de l’Homme, courtesy artist J. Blanedet. Behold:
Finally, a sauropod life restoration! J. Blanedet's 1886 Atlantosaurus poses with an elephant for scale. From Flammarion (1886).
Flammarion’s book is a landmark work for depictions of prehistoric life, containing a mix of old and new restorations that went some way to relieving the drought of new reconstructions in the late 19th century. Perhaps reflecting Flammarion’s outsider position from sauropod research, his choice of sauropod was not something well-known like Cetiosaurus, Camarasaurus or Brontosaurus, but the obscure “Atlantosaurus”. This was one of the first discovered Morisson Formation sauropods but, on account of its scrappy remains, it was on its way to becoming a historic footnote in 1886. Today, Atlantosaurus is generally considered a nomen dubium. Obscure taxon choice aside, here, finally, was a sauropod in the flesh. And, all things considered, the reconstruction was pretty good. Marsh’s 1883 Brontosaurus reconstruction was featured in the same book and its shared DNA with the Atlantosaurus illustration is obvious. In the accompanying text, Flammarion describes several sauropods in essentially accurate ways: as gigantic, long-necked animals with small heads, of herbivorous character, and as denizens of terra firma, not lakes and swamps. This was a pretty progressive take on sauropods that built on ideas expressed by Phillips, and they stand in contrast with sauropods’ fast approaching 20th-century relegation to semi-aquatic life.
In a different universe, Flammarion’s Atlantosaurus and other novel reconstructions were the start of a new wave of palaeoartworks based on updated science and newly discovered species. Alas, in our universe, the new artworks in Flammarion’s book made little impression on palaeoart development, and didn’t shake older takes on extinct animals from their foothold in 19th century palaeoart. Even as the 20th century loomed, Hawkinsian, Kuwassegian and Copeian dinosaurs were still populating artworks of Deep Time. From a historic perspective, Flammarion’s Atlantosaurus is more of an Easter egg than the moment sauropod palaeoart truly arrived.
Joseph Smit's 1892 Brontosaurus, from Henry Hutchinson's Extinct Monsters: A Popular Account of Some of the Larger Forms of Ancient Animal Life. Although not as influential or well known today as the sauropod artworks of Charles Knight, the success of Hutchinson's book would have made this the first life restoration seen by many people, certainly in the English speaking world.
It was only as the sandgrains in the 19th century hourglass ran dry that a collective epiphany about sauropods struck the minds of museum developers, artists and book authors around the world. Finally, after more than a half-century of avoiding sauropod palaeoart, flesh restorations of long-necked dinosaurs entered the mainstream during the 1890s. But, again, it was not America, with its embarrassment of sauropod fossils, that instigated this. Rather, it was a revived British interest in palaeoart that championed sauropods, with one book, in particular, cresting a new wave of new palaeoartistic reconstructions: Henry Neville Hutchinson’s 1892 ;Extinct Monsters: A Popular Account of Some of the Larger Forms of Ancient Animal Life. This featured illustrations by the Danish artist Joseph Smit based on Marsh's skeletal diagrams and the results were so successful that Extinct Monsters was reprinted several times over the next two decades. It was also eventually repackaged with another Hutchinson book, Creatures of Other Days. Smit illustrated both Diplodocus and Brontosaurus in these works, based on Marsh's groundwork, establishing their canonical presence in palaeoart from then onwards.
One of the most iconic palaeoart images of the late 19th century: Charles Knight's 1897 swamp-bound Brontosaurus and grazing Diplodocus. Knight's affiliation with the American Museum of Natural History and the publicity-hungry Henry Osborn brought palaeoart into a new era where dinosaurs would become increasingly dominant art subjects, and sauropods would become superstars.
Shortly after, American palaeoart finally woke up to the splendour of sauropods. The American Museum of Natural History, under the supervision of Henry Osborn, had identified the power of palaeoart as an educational, promotional and commercial tool and realised the role dinosaurs could play in this campaign. The result were sauropod illustrations by Charles Knight, which surely rank as the most famous of all early sauropod artworks. Knight’s iconic 1897 Brontosaurus and Diplodocus in a swamp, produced with direction from Osborn, was among the first of these works, but a lesser-known sketch of a snorkelling Amphicoelias was produced around the same time. The latter is notable for being produced under guidance from Cope, who sketched a rough version for Knight to replicate. Cope’s choice to depict the fragmentarily known Amphicoelias, which he named in 1878, over a better-known Marsh species is surely an instance of their old rivalry dying hard. Many Knight images of sauropods would follow in the next few decades.
In the last year of his life, Edward Drinker Cope tutored Charles Knight in the anatomy of extinct animals so as to help Knight better understand his palaeoart subjects. Among the images they worked on together was this 1897 scene of Amphicoelias, which Cope sketched out for Knight to replicate. Note the interesting stripes and spotting used here. Although now standard in palaeoartworks, such intricate patterning was rare in 19th century palaeoart.
And it’s here, at the turn of the 20th century, that our story picks up with more familiar beats. After their slow adoption into palaeoart and popularised palaeontology, sauropods quickly appeared everywhere dinosaurs were mentioned. American museums put sauropods front and centre of their galleries and, courtesy of Andrew Carnegie, many museums around the world soon sported Diplodocus in their dinosaur halls. Gertie the Dinosaur and the 1925 adaption of The Lost World, where Brontosaurus pioneered cinematic monsters smashing cities for our amusement, made sauropods the focus of early dinosaur films. Life-sized models of sauropods appeared in Europe by 1910 and then in the United States and Russia in the 1930s. Around all this, artists like Knight, Smit, Heinrich Harder and Alice Woodward cemented sauropods into the mainstream palaeoart canon. Surely helped by their palaeoartistic popularity, sauropods had become cultural icons almost overnight.
The story of sauropods entering palaeoart canon is more than a tale of 19th century attitudes to reconstructing extinct animals: it’s also a case study in the vagaries of science communication and gatekeeping. We often discuss why certain facets of science, like dinosaurs, are so popular, while others are not. How much of this reflects the inexplicable, innate interestingness of a topic, and how much of it is manufactured? Many of us would agree that sauropods are some of the most fascinating and spectacular animals to have ever lived but, even when their anatomy was well-realised, this was not enough for mainstream culture to adopt them passively. Even artists and scientists who knew about them were not falling over themselves to restore and promote them, and it really wasn’t until individuals at the turn of the 20th century wanted, or needed, to promote sauropods that they began their journey towards being palaeontological icons. I’ve argued in the past that dinosaurs have a certain fundamental appeal that draws us to them, and that might be true, but stories like this show that our awareness and access to spectacular, easy-sell science, such as that of the biggest dinosaurs to have ever existed, is managed by a privileged few.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Bramwell, V., & Peck, R. M. (2008). All in the bones: a biography of Benjamin Waterhouse Hawkins. Academy of Natural Sciences.
Dodson, P. (1998). The horned dinosaurs: a natural history. Princeton University Press.
Flammarion, C. (1886). Le monde avant la création de l'homme: origines de la terre, origines de la vie, origines de l'humanité. C. Marpon et E. Flammarion.
Hallett, M., & Wedel, M. J. (2016). The sauropod dinosaurs: life in the age of giants. JHU Press.
Nieuwland, I. (2019). American Dinosaur Abroad: A Cultural History of Carnegie's Plaster Diplodocus. University of Pittsburgh Press.
Phillips, J. (1871). Geology of Oxford and the Valley of the Thames. Clarendon Press.
Secord, J. A. (2004). Monsters at the crystal palace. In: de Chadarevian, S, & Hopwood, N. (eds). Models: the third dimension of science, Stanford University Press. 138-69.
Taylor, M. P. (2010). Sauropod dinosaur research: a historical review. Geological Society, London, Special Publications, 343(1), 361-386.
That most obscure theropod taxon Tyrannosaurus chews on bones on the front cover of Emily Willhoughby's new book, Drawing and Painting Dinosaurs: the latest entry into the growing literature dedicated to palaeoartistry. Can we talk about how nice that tyrannosaur knee is? From the Crowood Press website.
The last decade or so has seen the arrival of several notable palaeoart books, articles and book chapters that showcase the works and voices of palaeoart practitioners past and present, such that it seems we’re in a particularly rich literary era for this specialised artform. Among these works are a slowly growing number dedicated to palaeoart methodology: the hows, whats and whys of restoring the life appearance of extinct organisms from fossil remains. In late October of this year, another book in this vein arrived to add to your palaeoart library: Emily Willoughby’s Drawing and Painting Dinosaurs.
I was fortunate enough to be sent a review copy of this new book by Crowood Press, who you may know from my own The Palaeoartist’s Handbook (2018) and the upcoming Witton and Michel volume The Art and Science of the Crystal Palace Dinosaurs(coming May 2022, vintage palaeoart fans!). Much as I wanted to write about Emily's book, my involvement with Crowood presents a conflict of interest to presenting any thoughts I may have, and the fact that Emily kindly contributed artwork to The Palaeoartist’s Handbook only complicates matters further. But in wanting to do something to promote what I think is a useful, welcome addition to our collective palaeoart bookshelf, I reached out to Emily to see if she’d agree to an interview about creating Drawing and Painting Dinosaurs. As you’ll have guessed by now, Emily kindly agreed to answer my questions and the full interview is below.
But before we get to that, we should give a quick introduction to the book in question. Split into eight chapters and two appendices, Drawing and Painting Dinosaurs is a good-sized (280 x 220 mm, 176 pages), well-produced and affordable (RRP £18.99) softback that covers the basics of the palaeoart process as well as reconstruction approaches to several dinosaur groups. Chapters 1-4 cover the basics of palaeoartistry, from restoring anatomy to recreating environments, chapters 5-6 cover restoring pennaraptorans, tyrannosauroids, ornithischians and sauropods, and chapter 8 uses the evolution of feathers as a case study for palaeoartistic prediction. Emily’s qualifications to write such a book, of course, are in no doubt. She is one of the leading palaeoartists of modern times and has been particularly influential in the field of restoring feathered dinosaurs - especially dromaeosaurs. Readers will surely be familiar with Emily’s takes on these animals from her online presence (website, Facebook, Twitter), press release artworks, museum exhibitions and inclusion in landmark palaeoart collections (e.g. Titan Books’ Dinosaur Art II). Even if you’ve been living under a palaeoart-impervious rock for the last decade and somehow missed Emily's stuff, the simple fact is that anyone who can draw and paint dinosaurs like this…
...is clearly someone to pay attention to when they're offering pointers and advice on restoring fossil organisms.
Drawing and Painting Dinosaurs is packed with illustrations - over 250, according to the back cover. Many of them are new (at least, I didn't recognise them from other sources) and it's fun seeing Emily take on taxa we've rarely seen her restore before - giant dinosaur herbivores, big carnivores and so on. You don't need to read a word to realise that this is a must-buy for palaeoart fans: simply having page after page of Willoughby palaeoartworks on your bookshelf is worth the cover price alone. Emily’s world-leading reputation is well-earned for her attention to detail, technical excellence and eye for composition. Eschewing the open plains, giant animals and big skies that have been a staple of dinosaur palaeoart for generations, Emily’s artwork is often more intimate, frequently set in densely forested habitats with fallen logs, patchworks of light and colour, and delicate foliage. Her restorations are not only enormously charismatic but also grounded in observations of modern species, good knowledge of animal behaviour, and an appreciation for real natural spaces. Her combination of skills and approaches makes her Mesozoic dinosaur artwork fantastic to look at and also eminently believable. It's difficult not to think her paintings (especially her more recent and detailed pieces) were not drawn from scenes witnessed with her own eyes. We might not know for certain what Mesozoic dinosaurs looked like or how they behaved, but Emily’s artwork is surely in the right ballpark.
Fortunately for those of us secretly plotting to steal Emily’s artistic essence who’d like to learn to restore dinosaurs with that Willoughby touch, Drawing and Painting Dinosaurs has a greater emphasis on artistry and technique than we’re seen in most other palaeoart guides published to date. The closest comparisons I can think of are Douglas Henderson's palaeoart chapters in the first two Complete Dinosaur books but, with a full book of her own, Emily obviously has a lot more opportunity to discuss her craft. She outlines several methods used in creating her artworks such as painting from models, drawing over articulated fossils, and finding inspiration among real environments, while also giving pointers on matters such as composition, traditional painting techniques and finding basic forms within dinosaur bodies. This approach, combined with her patient, clearly-written text, will make the book especially useful to non-specialists. Technical terms are used here and there, of course (it’s basically impossible to write at length about palaeoart theory without some jargon) but in-text explanations and a glossary make the introduction of such terminology a learning experience, not a barrier to understanding. This is not to imply that the book is just for beginners, of course: there are plenty of useful ideas and takes on dinosaur palaeobiology that will be invaluable to artists of all levels. There were certainly some facts, perspectives and methods that were new to me, for whatever that's worth.
A sample page from Drawing and Painting Dinosaurs, from the Crowood Press website. This page features Emily's drawover of the Mei long holotype, a technique that helps artists not only understand the anatomy of their subjects but also appreciate fossil specimens as the remains of individuals, not as mere scientific concepts. Seeing fossils as the remains of specific creatures forms one philosophical core of the book.
And speaking of the text, I found Drawing and Painting Dinosaurs pleasant to read for its informative, slightly conversational tone. It presents a unique voice in palaeoart discourse, neither written with the disembodied neutrality of a scientist nor with overconfidence about her preferred interpretations of the past. There's an obvious respect for the work and insight provided by scientists but also plenty of informed personal contemplation and opinion on matters of reconstructing anatomy and ancient environments. The book is sprinkled with reflections on specific artworks and projects that give a sense of the enjoyments and frustrations of the palaeoart experience, such as including having artwork dating within days of its completion, the intrigue of reworking a familiar taxon with new data, and the thrill of restoring a newly discovered species. It not only reassures us that Emily is experienced at the trade she’s teaching but gives the book a sense of personality. We're also given insights into how Emily views the past, and her obvious connection to her fossil subjects stands out as something I've not seen expressed in palaeoart literature before. Emily reminds us that fossils are not mere geological phenomena or abstract concepts like species, but the petrified tissues of individuals that lived and died for us to discover millions of years later. It's a sobering, thoughtful take on palaeoart that establishes a personal connection between artist and their extinct subject matter across Deep Time.
One inescapable feature of Drawing and Painting Dinosaurs is its strong focus on dromaeosaurs and related, fully-feathered theropods - especially Deinonychus - for both artistic and case study subjects. These animals really do take centre stage - something like 80% of the artwork features dromaeosaurs or similar dinosaurs - and they serve as go-to species for demonstrating palaeoart principles throughout most of the book. Tyrannosauroids, sauropods and all ornithischians also feature in their own discrete chapters, but Drawing and Painting Dinosaurs is undeniably a show driven by feathered dinosaurs. I feel this is the only aspect of the book that might prove divisive, especially if readers are expecting a more general guide to the life appearance of dinosaur groups. There is value, however, in this doubling down on pennaraptorans. Writing any text like Drawing and Painting Dinosaurs always boils down to a question of breadth vs. depth and, in choosing the former, the book sacrifices some use as a general reference for demonstrating how a deep understanding of a fossil group can enhance our palaeoartistry. Knowing every species, every trace fossil, and every specimen of a clade allows for especially informed and nuanced palaeoartistic approaches, and that’s strongly evident in Emily’s pennaraptoran dinosaur art. For less experienced artists, the amount of information she demonstrates can be transferred from fossils to palaeoartworks may be surprising, and this would not have been so obvious had Emily discussed more clades in less detail. Seeing what can be done with dromaeosaurs and their relatives provides an impetus to learn about our favourite subjects in just as much depth, for which readers are given the right guidance for what to research and where to find it.
Anyway... this isn't meant to be a review, and here I am writing everything I like about the book. That should give you a flavour of what the book is all about and, as you can tell, I have nice things to say about it. With claxons blazing for those conflicts of interest mentioned above, I recommend anyone interested in palaeoart check it out. But we're not here for my thoughts: let's move on to what we’re actually here to read - Emily’s insights into how the book came to be, the original plans for the project, how she creates her artworks and even some free tips for us budding artists. Huge thanks to Emily for agreeing to this interview, and I hope it inspires you to put Drawing and Painting Dinosaurs on your Christmas list, if it’s not there already.
--
MW. It’s felt like a book such as Drawing and Painting Dinosaurs was going to happen eventually: a professional, world-leading palaeoartist imparts their experience and knowledge about illustrating the most popular and in-demand palaeoart subjects. I’ve certainly been around pub tables where artists have discussed it. What made you step up to the plate and think “yep, I’ll take that project on”
EW. It had been a dream of mine to publish a book like this someday, but my experience thus far with bringing book proposals to publishers and agents had been largely disappointing. For my first book, God’s Word or Human Reason?, I and my coauthor sent a total of about a hundred query letters to agents without a single positive response. So when Crowood Press reached out to me and asked if I’d be interested in such a project, there was no hesitation on my part!
Your text has an especially patient, welcoming quality that carefully explains a lot of information for newbies, encourages readers to draw along as they read, and there are several step-by-step illustration guides. It contrasts with what I’ve come to expect from palaeoart guides, which can slide into machine-gunning anatomical facts and interpretations about extinct animals at the reader. Was this simply your intuitive approach, or something you deliberately crafted when writing your book?
You were actually a pretty big inspiration for the approach I decided to take in this book. Since Crowood also published your wonderful The Palaeoartist’s Handbook, I knew from the outset that I needed to take an approach that would not overlap too much in content and style with your own. I played with a lot of different ideas early on — including, for example, a separate chapter on each medium I typically work in (gouache, digital, pencil, and so on), but felt that approach was too “arty” and not “sciency” enough. Ultimately I wanted to create something that seamlessly entwined the scientific and epistemological bases of paleontology with the hands-on artistic techniques, and this led to a structure that attempted to educate both professional and lay readers by providing context-driven examples.
This book was an enormous education to me in how damned difficult it is to craft a coherent structure of an entirely new book without much outside feedback. The first month or so of its creation was dedicated entirely to what seemed like endless deliberation, revision, and hand-wringing over the structure, approach and focus. I think the decision I went with turned out decently enough, but I still have a lot to learn in this respect.
Drawing and Painting Dinosaurs has a huge number of images - over 250 - and you created virtually all of them yourself. How much of that was new artwork for this book, and how long did it take to produce? A lot of them seem very recent - were you basically painting 24/7 until the book was finished?
I would estimate that over half of the illustrations are ones that were created specifically for this book, including a large selection of less time-intensive diagrams and sketches. Of the illustrations that were previously completed, many of them were progress shots or thumbnails that haven’t been put to good use until now (I knew there might a good reason someday for me to save progress shots of long-ago artworks!). A handful were also from various projects over the years that never managed to see completion for one reason or another, and some were finished up from early works-in-progress that were subsequently abandoned.
I had about a year total to complete the book from start to finish. I’m still kind of dumbfounded that I managed to get the book together during the same year in which I wrote and defended my Ph.D. dissertation! I have no doubt that’s something I’ll look back on in my twilight years and think “how on earth did I ever have the energy!”
We get to revisit some of your older artworks in Drawing and Painting Dinosaurs which have a slightly different style and mood to your more recent work. Thinking specifically about palaeoart-specific skills, how has your art and approach developed since their creation?
I still have a long way to go, but I like to think that my work has improved tremendously since I first started doing serious paleoart. For one thing, I’ve learned to enjoy reconstructing taxa I’m relatively unfamiliar with, whereas I started out painting feathered theropods and little else. While the new book still largely features dromaeosaurs and their kin, I had a lot more fun than I expected to have on the sauropods, tyrannosauroids, and various ornithischians that were included. For another thing, I’ve expanded my artwork to a variety of media I never worked in regularly early on—oil paint, gouache, acrylics, and graphite.
I’ve also become more comfortable in testing out new ideas and compositions. Recently I’ve been putting more effort into the environment and setting, which I used to think of as a chore that took away from the fun of painting the dinosaurs themselves. But once it starts becoming fun to paint environments, it also becomes easier. My experience is that the single largest improvement in my artwork isn’t so much in a piece’s overall quality per se, but in learning to work more efficiently—better quality per unit time.
As anyone familiar with your portfolio might expect, Drawing and Painting Dinosaurs has a very strong focus on dromaeosaurs and their relatives, especially Deinonychus. What’s the draw of these dinosaurs over sauropods, ornithischians or more rootward theropods?
It’s no secret that my biggest focus in paleoart has always been dromaeosaurs and other feathered coelurosaurs—I’m a feather-fancier, it’s true. Although I did enjoy branching out a bit for this book, dromaeosaurs remain the group of dinosaurs that I find most captivating and arresting. I suppose the reason for this is that feathered dinosaurs were responsible for my introduction to paleoart after a lifetime of obsessive interest in birds and evolution in general. In the early 2000s, I recall reading articles about some of the exquisitely preserved Liaoning fossils, including Sinorthithosaurus, Sinosauropteryx and especially Microraptor. When I first saw a photograph of the fossil of Microraptor in 2003, I was utterly fascinated, and it caused the realization that birds were dinosaurs to register new and profound understanding. The rest is history!
There’s an emotional component to your book that is unusual for discussions about the technicalities of palaeoart. You write about the personal affinities and emotions you’ve developed for certain subjects, and that you regard fossils as not just specimens of not only long-extinct animals, but as long-deceased individuals. An ode to extinct animals, the poem Not Forgotten by Jonathan Kane, ends chapter 1, and you describe Deinonychus as possibly being “the most beautiful animal that ever lived”. There’s a clear attachment to many of your extinct subjects - how much of an influence does this have on your artwork, and do you feel the same about all extinct animals? Or are some subjects “just a job”?
I do think that many natural subjects are beautiful, and it’s hard to not express this when trying to communicate that beauty in my own art. Although I used to think of some natural subjects as “just a job” (especially environments and flora), I feel less that way over time—all natural subjects are beautiful and fascinating, though of course I have my own attachments and biases. Moreover, though, I think that communicating this passion to the public is important to encourage people to think of dinosaurs as real animals, and to cultivate a sense of awe and respect that I feel extinct animals deserve.
You detail both digital and traditional painting techniques in your book. Do you have a preferred medium?
Part of what I enjoy about working in different media is that it’s harder for me to become bored and frustrated. Sometimes I get annoyed at how cluttered and messy my study gets when I work in gouache and (especially) oils for a while, so I switch back to working digitally for a while. Then I may start getting annoyed at how often Photoshop crashes or how Procreate constrains layers and resolution so harshly, so back to gouache it is! Learning and practicing new media is always interesting and keeps me engaged. I need variety. Currently, my favourite medium to work in is Procreate—I love how easy it is to blend, and I can take my iPad to meetings and work. Gouache has always been my traditional medium of choice, but that may change as I work more in acrylics and oils.
I’m also curious to know if you’ve attempted to update traditional paintings when new science forces us to revise older reconstructions.
The one piece of traditional art I spent a lot of time repainting was my 2010 painting of Anchiornis, which I talk about in the book. New to oils at the time, I painstakingly painted a mottled brown and white birdlike critter dashing through a lush green jungle. It was literally days after I was finally satisfied with its completion that the first color study was published, showing that Anchiornis was more likely to be black, grey and red than brown and white. I could have justified the piece as a juvenile, subspecies, or female, but instead I repainted the animal and adjusted the background to the new contrast it required. I’m glad I did, and I sometimes do include minor updates to older pieces, but usually I prefer composing a new piece altogether when new data comes to light.
The palaeoart community has long been dominated by males, both in terms of practitioners and fans, such that Drawing and Painting Dinosaurs is among the first, if not the first, palaeoart guide written by a female author and artist. With so much discussion around the lack of diversity in palaeontology, your book feels like an important milestone in diversifying the voices shaping palaeoart. Was this something you were conscious of when creating your book
I don’t think it occurred to me at the time that I was the first, given that there aren’t many paleoart guides out there to begin with. But I’m honoured to represent that milestone if so, and I hope the example may encourage others. I’ve already gotten a few emails and even an interview for a project from a few school-aged girls who want to become palaeoartists, which is frankly one of the very most rewarding things about working in this field to me.
You cover a heck of a lot of ideas and techniques in your book, from finding inspiration among natural settings to building models, to constraining colour patterns using living animals to modifying modern plant leaves to resemble those of the past… and much more. Out of this extensive toolbox, are there any that you regard as especially essential - the one or two that, whatever your approach to palaeoart is, we should all be doing?
Remember that prehistoric subjects were living things, and look to other living things (their anatomy, their ecology and evolution, their behaviour, the way they move and interact in their environment, and so on) for inspiration to capture that sense of lifey-ness that lifeless things are meant to have in paleoart. Although many prehistoric organisms were quite unlike anything alive today, they all share an important thing: they evolved to live and move in their environments. I think that this single injunction can go a long way towards facilitating accurate, interesting, and unique paleoart.
It’s not always wise to ask about the Next Big Project so soon after finishing one, but are there any big Willoughby projects we should look out for soon? Emily Willoughby’s SuperBig Dulux Coffee Table Book of Dromaeosaur Art has a nice ring to it...
So glad that you asked! I am about halfway through a new project: Marks on Time, an illustrated anthology of natural history poetry written by my coauthor Jonathan. This will be a collection of 25 poems about evolution, dinosaurs, and human nature, accompanied by about 40 full-colour and brand-new illustrations. If any publishing agents happen to be reading this, please drop me an email. ;)
--
Thanks again to Emily for her interview responses, and I wish her all the success her book deserves. Drawing and Painting Dinosaursis available now from Crowood Press, priced £18.99, as well as from all the usual book retailers.
Enjoy these insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
The story of how dinosaurs were resurrected for cinema in the early 20th century tends to focus on the visual components of these technical feats: the 2D animation and stop-motion technology that made long-extinct reptiles move again. But alongside making dinosaurs walk and run, cinematic dinosaurs also gave them a voice, imagining their snarls, barks, screeches and roars. Perhaps the first film to do this (or certainly the first mainstream, widely-seen film, at any rate) was 1933’s King Kong, where a charging Stegosaurus might take the title of first vocalising dinosaur in cinematic history:
The crew of the Venture encounter a Stegosaurus: a famous scene from 1933's King Kong and, potentially, the first dinosaur cinema audiences ever heard vocalise. From Youtube's Prehistoric Classics.
The same film would give us plenty of other prehistoric animal action and noise, but by far the most famous is surely the snarling, screeching Tyrannosaurus:
Tyrannosaurus takes on King Kong in er, King Kong (1933). We've all seen it a million times, but go on, watch it again: you know you want to. Note the screechy tyrannosaur noises, presumably to contrast with Kong's throaty roars. From Youtube's Movieclips.
Over the last century, cinematic dinosaur appearance and the technologies used to depict them have changed dramatically but one thing has remained the same: dinosaurs are as loud and noisy as ever. We’ve even seen the development of conventions and tropes around what dinosaurs sounded like. Sauropods are often given haunting, humpback whale-like songs and dromaeosaurs are frequently assigned aggressive, high-pitched snorts, snarls and crackles. And, of course, big theropods — especially Tyrannosaurus — invariably have deep, bellowing roars. It’s no exaggeration to say that the Jurassic Park Tyrannosaurus roar is as iconic and recognisable as its now-famous (maybe infamous?) design.
The desire to depict talkative, raucous dinosaurs has long transcended media capable of conveying sound. Even movie dinosaurs were noisy and boisterous before we had the technology to make them truly roar on screen. The 1925 silent film The Lost World features plenty of roaring, snarling and bellowing stop-motion dinosaurs framed by director Harry O. Hoyt in dramatic close-up. Even in silence, the intent of these shots is obvious, and we simply have to imagine their vocalisations ourselves. Fantasia’s famous 1940 Rite of Spring sequence performed a similar trick for artistic reasons, juxtaposing a roaring Tyrannosaurus against booming segments of Igor Stravinsky’s famous composition. Static, traditional palaeoart also has a strong emphasis on animal vocalisations. Peruse any gallery of prehistoric animal restorations (such as this, at my new website!) and we inevitably find heaps of artworks showing grunting, chirping, screaming dinosaurs. Viewed objectively, it is a little strange that we focus so much on this behaviour in our artwork. Why don’t we render more non-acoustic behaviours that are arguably better suited to a totally visual medium? And furthermore, why draw so much attention to an aspect of dinosaur behaviour we don't know much about? Subconsciously, we just can’t get away from the call — pun not intended — of depicting extinct animals vocalising.
An Archaeopteryx siemensii perches on driftwood, opens its mouth and calls out... what, exactly? We palaeoartists can't resist drawing animals posed mid-vocalisation, despite our lack of knowledge about extinct animal sound production.
Clearly, we've collectively decided that prehistoric reptiles were vocal, noisy species, and this is understandable. It is, after all, what we experience around us today. Our world is full of singing, calling birds and barking, bellowing mammals. Whales sing, lions roar, and frogs croak. It stands to reason, then, that dinosaurs would be just as vociferous, and that a Jurassic or Cretaceous dawn would be full of strange, wondrous hoots, bellows, chirrups and songs that we can only imagine. I am, of course, leading up to a weighty “however”. What if our assumption of noisy, vociferous dinosaurs is simply... wrong?
This is, of course, a very strong accusation, especially because we can say very little definitively about dinosaur vocalisation owing to our lack of fossilised dinosaur throat tissues and vocal organs. The preservation of such anatomies among Mesozoic dinosaurs is not impossible, these having been found in Vegavis iaai, a Mesozoic bird that lived in Antarctica 69-66 million years ago (Clarke et al. 2016); but this remains an exceptional occurrence: Vegavis is the only Mesozoic dinosaur known with preserved vocalisation anatomy. We can, however, use fossils and data from extant reptiles and birds to make predictions about dinosaur vocal ability, and several researchers have attempted this (e.g. Weishampel 1981; Senter 2008; Brazaitis and Watanabe 2011; Clarke et al. 2016; Reide et al. 2016). Among the more famous examples of such works is Phil Senter’s 2008 Voices of the past: a review of Paleozoic and Mesozoic animal sounds, a synthesis of what we know of sound production among ancient animals. In his section on birds and their ancestors, Senter makes the bold suggestion that non-avian dinosaurs may have been — yikes — entirely non-vocal (Senter 2008). In other words, this posits that dinosaurs may have not only been much quieter than their pop culture counterparts, but actually reliant on non-vocal acoustics when they wanted to communicate audibly. This notion — which I’m calling the "silent dinosaur hypothesis” — gained a fair bit of discussion online when first published and still crops up in modern conversations about dinosaur behaviour. But how does it hold up over a decade on, and did it ever have a sound basis to begin with?
Non-vocal (or, at least, closed-mouth) interaction between male and female Ceratosaurus nasicornis. Were dinosaurs limited to posturing and other means of display for their communication with one another? Some hypotheses suggest so.
To explore this further, it will help to outline what non-vocal animal acoustics are. We animals make noise all the time simply by existing and going about our lives. These ‘passive’ noises are classed as non-vocal acoustics. They include sounds that come from acts like breathing, forcing air around your throat tissues, and hitting or rubbing body parts against each other or external objects. Many animal species exploit these phenomena to make deliberate, structured sounds for communication. A hiss, for example, is little more than forcefully expelling air through our throats and mouths. A snort is much the same, except using our noses. We can also purposefully slap or rub body parts together or against the ground, water or vegetation to create loud noises. Some species have developed special anatomy purely to create non-vocal sounds, with the most obvious example being rattlesnake tails. Non-vocal acoustics are everywhere once we start noticing them, and Senter (2008) argued that they may have been the only sounds made by dinosaurs. It's incontrovertible that these are the only noises we can be confident that dinosaurs made because they can be generated regardless of vocal capability. Whatever other noises dinosaurs created, we know that they could hiss, snort, stamp their feet and so on, and living diapsids show that such behaviours are used as communication strategies among extant dinosaur relatives. Non-vocal acoustics are also perfectly compatible with the large noses and crests that may have acted as resonating chambers in certain dinosaurs, too (e.g. Weishempel 1981; Witmer and Ridgely 2009). We know, for instance, that some non-vocal snake species use resonating cavities in their throats to turn hisses into growls (Young 1991; see an example here of the slightly terrifying noises from of a king cobra).
So, yes, non-vocal acoustics make a lot of sense for dinosaurs — documentary makers, take note. But Senter's (2008) accusation is that dinosaurs could only make non-vocal sounds, and that requires us to consider 'true' vocalisations: the sounds animals make by forcing air through their vocal organs. Mammals, amphibians and non-avian reptiles use a larynx for this purpose, while birds have their own, unique voice organ: the syrinx. It’s the evolution of this structure that prompted Senter’s suggestion of non-vocal dinosaurs. Unlike the larynx, which is situated at the top of the throat, the syrinx is located at the base of the trachea where the airway forks into the lungs. It also works in a different way to a larynx. Rather than passing air over vocal folds, the syrinx generates sound from the airway walls themselves. Rushing air from the lungs flutters these membranes and associated cartilages in a manner that produces sound, and the location of the syrinx at the fork where the trachea becomes a pair of bronchial tubes allows for especially complex vocalisations: each bronchial component can vibrate asymmetrically, making two sounds at once. To keep their airways open, avian syrinxes are reinforced with well-mineralised cartilaginous rings. In modern birds, it seems that syrinxes can also only function with assistance from a clavicular air sac (Senter 2008), although experiments indicate vocalisation without this structure may be possible (Clarke et al. 2016).
Thanks to fossils of Vegavis, we can be confident that extinct duck and goose relatives like Conflicto antarcticus — a Palaeogene species from Antarctica shown here — were capable of making honking, goose-like sounds. But we have very little direct evidence for the sort of noises more rootward dinosaurs were able to make.
The reinforced structure of the syrinx and its possible association with an air sac means that, unlike larynxes, they have some geologically detectable elements. In theory, this allows us to gauge roughly when, and in which lineages, they evolved even if fossil syrinxes themselves are rare. We can search for evidence of the clavicular air sac pneumatising the bones of the pectoral girdle and forelimb, as well as fossils of those reinforcing, mineralised rings holding the syrinx open. Although soft-tissue in nature, these structures are found in Cenozoic bird fossils (Clarke et al. 2016), so they evidently have decent enough fossilisation potential in the right circumstances.
The results of such searches have come back without much to report, however. Even in well-preserved Mesozoic dinosaurs, we find no consistent evidence for clavicular air sacs outside of the ornithothoracines (the group of avialans that includes enantiornithines and crown birds, but see Senter 2008; Wedel 2009 for a few exceptions) and not a single mineralised airway has been discovered in a non-avian dinosaur (Senter 2008; Clarke et al. 2016). This suggests that the syrinx was developed very late in dinosaur evolution, perhaps not even being present in feathered, otherwise extremely-bird-like dinosaurs (Clarke et al. 2016; Kingsley et al. 2018). We should not assume, of course, that the avian syrinx sprang into existence fully-formed — surely it had to develop via intermediary ‘proto-syrinx’ structures first (Kingsley et al. 2018) — but we don’t know what that structure was nor what features might evidence its existence. With our present dataset, all we can say is that the avian syrinx as we know it probably wasn’t present in most non-bird dinosaurs. It follows that if dinosaurs did not have a syrinx, they were probably incapable of making the rich, complex noises of modern birds.
And this is where things get especially interesting. OK, so dinosaurs weren't singing like passerines, but most reptiles have a larynx, and we can be pretty certain that dinosaurs did too. So Senter must be wrong, right? Dinosaurs merely vocalised like modern reptiles: case closed. Well, not necessarily, because we don't know if the dinosaur larynx was functional. Many lizard larynxes lack vocal folds and thus cannot vocalise, and opinions differ on whether their vocal abilities were independently lost from a vocal reptilian ancestor (e.g. Kingsley et al. 2018) or convergently gained from a historically silent one (e.g. Russel and Bauer 2021). Furthermore, birds also have a larynx, but it's also non-functional. This leaves dinosaurs evolutionarily bracketed by crocodylians (with a functioning larynx) and birds (with a non-functioning larynx), creating ambiguity about the ancestral state of dinosaur vocal organs. The ancestral acoustic capabilities of other reptiles is thus very important to determining what the original state of archosaur vocalisation was. There are two possible models (Kingsley et al. 2018): perhaps archosaurs were ancestrally silent, with crocodylians and birds to developing functional vocal organs independently of one another; or they were vocal, with birds augmenting and/or replacing the larynx for an unknown reason late in dinosaur evolution. And this touches on another key question with bearing on dinosaur vocalisation: why did birds develop the syrinx at all? One possibility is that the syrinx evolved in response to having lost, or having never developed, a vocal organ in the first place (Kingsley et al. 2018), a scenario implying that at least some theropods, if not all dinosaurs, went through a silent phase in their evolutionary history. The bottom line is that there's still a lot to learn about the evolution of reptile vocalisation, and there are reasonable, entirely plausible models that align with Senter’s (2008) proposal that dinosaurs were non-vocal (below).
A handy graphic showing two competing models of syrinx evolution, from Kingsley et al. 2018. This assumes that reptiles were ancestrally vocal, but this doesn't change considerations of syrinx evolution too much. Essentially, we have two options: birds evolved a syrinx alongside a functioning larynx, or the dinosaur larynx wasn't functional, and the syrinx evolved as a novel structure to exploit vocal communication. The latter model, of course, implies at least some non-vocal dinosaurs.
These ideas are, of course, very difficult to test without appropriate fossil data. Senter (2008) noted some support from non-vocal lizards using visual communication instead of aural, thus placing extra significance on the often extravagant display structures of dinosaurs. Might all those crests, horns, frills, fancy scales and elaborate feathers have evolved because dinosaurs were essentially mute, primarily visual communicators (Senter 2008)? A counterargument to this is that lizards communicate visually without such crests, horns and so on, but the concept of some dinosaurs using display structures to compensate for a lack of vocal capability is still an interesting idea.
But before we get carried away with all this, we should note that the silent dinosaur hypothesis is not the only model of archosaur acoustic evolution on the table. A case can be made that, whatever weirdness was going on with syrinx evolution, dinosaurs were still capable of making laryngeal sounds. It has been noted that birds and crocodylians share several similar vocal behaviours that implies inheritance from a shared, vocal ancestor (e.g. Brazaitis and Watanabe 2011; Clarke et al. 2016) and some models of reptile evolution posit that all reptiles were ancestrally vocal, implying a functioning larynx in Dinosauria (Kingsley et al. 2018). Such concepts predict that dinosaurs vocalised at least in relation to matters of territory and courting, as well as to communicate between parents and offspring (Clarke et al. 2016). It’s difficult, of course, to know what specific sounds were made, and this isn't just because larynxes rarely fossilise: it's also because reptilian vocal anatomy is just not as well studied as that of birds and mammals (e.g. Rittenhouse et al. 1998; Reide et al. 2015; Russel and Bauer 2021). Recent work has shown that, although most reptile vocalisations are relatively simple compared to those of frogs, mammals and birds, there is a lot of variation in larynx structure across reptile species, and that their vocal tissues and acoustic capabilities can be very sophisticated, sometimes competing with mammals and birds in complexity (Brazaitis and Watanabe 2011; Reide et al. 2015; Russel and Bauer 2021). Among the most developed reptile vocal capabilities are those of crocodylians, which include a repertoire of behaviourally-specific hisses, grunts, bellows, snorts and chirps (Garrick et al. 1978), and those of geckoes, which use a range of single and repetitive chirps for advertising and alarm purposes (Russel and Bauer 2021). Perhaps, assuming dinosaurs did have functional larynxes, they made similar sounds.
I'll take any excuse to link to videos of bellowing alligators. American alligators typically bellow in water, but — as this video shows — they perform a similar behaviour on land, too. Note the closed mouth and inflating neck tissues here, and read on. From Youtuber JadeAtema.
We should clarify that the comparisons made by some researchers between reptilian and mammalian larynxes does not necessarily imply that dinosaurs vocalised like mammals. The throat tissues of reptiles and mammals are quite different in that reptiles can inflate their neck tissues with air from their lungs, whereas mammalian throat cartilage and muscles prohibit this action (Reide et al. 2016). This equips diapsids with a distinct mechanism for loud, deep sound production: closed-mouth vocalisation. By closing their mouths to prevent air escape and pumping air into their necks, reptiles and birds can create resonating chambers which allow for much deeper, lower-frequency vocalisations than could be achieved with a 'standard' open mouth call. We might intuitively think of crocodylians employing this behaviour to create loud, awesome bellows (especially the American alligator, which is the champion of crocodylian bellowing - see Garrick et al. 1978 and video above) but this tactic is not just used by big, exotic species: the cooing of pigeons and the ‘a-woo’ of eider ducks are also closed-mouth vocalisations. These acoustics have developed repeatedly throughout archosaur evolution and may have been practised by the dinosaur-crocodylian ancestor (Reide et al. 2016), so it seems reasonable to imagine this behaviour being used by dinosaurs making especially loud, deep and far-reaching noises. This may have been especially so among large species as, in birds at least, closed-mouth vocalisations have mostly evolved among bigger-bodied lineages (Reide et al. 2016). Predictions of archosaur voice evolution do not suggest that all dinosaur vocalisation would be closed-mouth (Reide et al. 2016), but those of us interested in depicting dinosaurs making their loudest, most intimidating noises should consider closed-mouth behaviours more likely than the usual stereotype of cat-like roaring (indeed, the roaring ability of Panthera species is associated with an unusual throat and laryngeal configuration (see Weissengruber et al. 2002), so we shouldn’t regard it as a ‘typical’ noise for any extinct animal to make, especially a reptile).
But we're getting a little off-topic now: we're here to talk about silent dinosaurs, not booming ones. So let's wrap things up. To summarise, there are a few take-homes here. The first is that the general assertion that we know nothing about dinosaur vocalisation isn’t really true: we certainly don’t know much, but we’re not entirely devoid of intelligent comment, either. A lot of the papers cited in this post are available online and are well-worth reading if you want to know more about the topics discussed above. The second is that the silent dinosaur hypothesis is far from a done-deal, but it has a more legitimacy than we might first expect. It's not, despite its unorthodoxy, a crazy idea and actually fits some interpretations of dinosaur vocal evolution, even if we can't really tell how right or wrong it is at the moment. There are huge caveats around any model of dinosaur vocal evolution, of course, the most important being that our models are so poorly informed by fossil data that one new discovery could turn everything we’ve predicted on its head. And that leads to a third main point: whatever ideas of dinosaur vocalisation we think are correct, we should appreciate that they’re not much more than personal preferences at the moment. But that’s fine, and it's even liberating for artists and filmmakers. This uncertainty gives us a huge playground for depicting dinosaur behaviour in ways we haven’t considered before. What does a non-vocal Tyrannosaurus do instead of roaring when it wants to look impressive? How do sauropods communicate without singing? What noises did Mesozoic birds make before they developed the syrinx? We don’t know, but it’s sure a heck of a lot of fun to think about.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Brazaitis, P., & Watanabe, M. E. (2011). Crocodilian behaviour: a window to dinosaur behaviour?. Historical Biology, 23(01), 73-90.
Clarke, J. A., Chatterjee, S., Li, Z., Riede, T., Agnolin, F., Goller, F., ... & Novas, F. E. (2016). Fossil evidence of the avian vocal organ from the Mesozoic. Nature, 538(7626), 502-505.
Garrick, L. D., Lang, J. W., & Herzog, H. A. (1978). Social signals of adult American alligators. Bulletin of the AMNH; v. 160, article 3.
Kingsley, E. P., Eliason, C. M., Riede, T., Li, Z., Hiscock, T. W., Farnsworth, M., ... & Clarke, J. A. (2018). Identity and novelty in the avian syrinx. Proceedings of the National Academy of Sciences, 115(41), 10209-10217.
Riede, T., Li, Z., Tokuda, I. T., & Farmer, C. G. (2015). Functional morphology of the Alligator mississippiensis larynx with implications for vocal production. The Journal of experimental biology, 218(7), 991-998.
Riede, T., Eliason, C. M., Miller, E. H., Goller, F., & Clarke, J. A. (2016). Coos, booms, and hoots: The evolution of closed‐mouth vocal behavior in birds. Evolution, 70(8), 1734-1746.
Russell, A. P., & Bauer, A. M. (2021). Vocalization by extant nonavian reptiles: a synthetic overview of phonation and the vocal apparatus. The Anatomical Record, 304(7), 1478-1528.
Senter, P. (2008). Voices of the past: a review of Paleozoic and Mesozoic animal sounds. Historical Biology, 20(4), 255-287.
Weishampel, D. B. (1981). Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology, 7(2), 252-261.
Weissengruber, G. E., Forstenpointner, G., Peters, G., Kübber‐Heiss, A., & Fitch, W. T. (2002). Hyoid apparatus and pharynx in the lion (Panthera leo), jaguar (Panthera onca), tiger (Panthera tigris), cheetah (Acinonyx jubatus) and domestic cat (Felis silvestris f. catus). Journal of anatomy, 201(3), 195-209.
Witmer, L. M., & Ridgely, R. C. (2009). New insights into the brain, braincase, and ear region of tyrannosaurs (Dinosauria, Theropoda), with implications for sensory organization and behavior. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 292(9), 1266-1296.
Quetzalcoatlus lawsoni scavenges a juvenile Torosaurus, recently killed (judging from the decapitation) by a large tyrannosaur. We've been waiting for ages to learn more about this pterosaur following a near-50 year delay in the description of its remains, but we're finally there! So what have we learned about Quetzalcoatlus proportions and anatomy from this glut of new information? Read on...
Almost a full half-century after its discovery, December 2021 finally saw the publication of a suite of technical papers on one of the most famous pterosaurs of all: Quetzalcoatlus. An entire themed collection of Quetzalcoatlus articles have been bound together in a special Journal of Vertebrate Palaeontology memoir, and they’re all — happy days — open access. This is great news for everyone with an interest in this animal or pterosaurs in general, and it represents a much-needed turnaround of a long-standing embargo on the Quetzalcoatlus material. Several authors have been attached and let go from The Big Quetzalcoatlus Project since the early 1970s and those of us without access to its fossils have done what we can to understand it from snippets of information published here and there. The memoir team — who tackle Quetzalcoatlus anatomy, systematics, palaeoenvironment and functional morphology — have thus ended a frustrating half-century-long wait for more information on this hugely popular, much-loved pterosaur. I have no doubt that these new papers — particularly Brian Andres and Wann Langston Jr.’s super-detailed and extensive osteology (Andres and Langston 2021) — will turn Quetzalcoatlus into a modern cornerstone of pterosaur research. Essentially overnight, the memoir team have turned Quetzalcoatlus from an almost non-entity in the scientific literature to an embarrassment of pterosaurian riches, and I look forward to the many new insights their work will foster.
There’s much to say about the entire volume and if you’d like a full run-down you should check out Darren Naish’s overview at Tetrapod Zoology. Here, I want to focus on one specific paper: the overview of functional morphology written by Kevin Padian et al. (2021). Many readers will have seen details on this paper in the media because it’s the one that turns our piles of Quetzalcoatlus fossils into a living, breathing animal, opining on what it looked like and how it walked, flew, and foraged. In other words, the kind of stuff that many people want to know most about these awesome animals. It's the paper that's drawn most of my attention because of my own research interests in azhdarchids: chiefly their palaeoecology, functional morphology and reconstruction (see Witton and Naish 2008, 2013; Witton and Habib 2010; Naish and Witton 2017 for examples).
And to cut straight to the chase, I want to talk about this because I suspect Padian et al.'s paper is destined to be the most controversial of the memoir’s contributions. It contains a lot of ideas and opinions that will be classed as unusual, non-mainstream takes on pterosaur palaeobiology and while some are novel, others are resurrected from papers written by the senior author decades ago (e.g. Padian 1983a, b, 1988, 2003, 2008). I say 'resurrected' because some of the ideas in question have since been rebutted or struggled to gain wider acceptance among pterosaur workers, such that reading Padian et al. (2021) has a distinctly vintage feel, like it's been beamed in from 20 or 30 years ago. Specifically, I'm interested in their handling of Quetzalcoatlus proportions, ground posture, wing folding and flight mechanics because Padian et al. make some genuinely strange suggestions around these topics; Quetzalcoatlus is restricted to crouching poses, might have flown with its legs tucked under its body, had its principal wing membrane attached to its hip, and potentially took to the air with a bipedal leap.
Proof that Padian et al. is destined to be controversial is found in the paper itself, where we find in-text admissions that the authorship team could not agree on matters of launch mechanic, flight posture and wing membrane configuration (in all instances, palaeontologist Kevin Padian is stated as disagreeing with his coauthors, biomechanicist Jim Cunningham and palaeoartist John “All Yesterdays” Conway). On top of this, some presented data conflict with other parts of the memoir, and there are some potential outright errors that affect the reliability of the paper’s conclusions. It is, in detail, something of a wild read, and reaction to this paper is going to be mixed among pterosaur experts. Nevertheless, Padian et al. (2021) will also be a potential source for pterosaur reconstructions for the foreseeable future, and given that both the paper and associated press coverage are publicly accessible, I think it's right to have some responses from pterosaur workers online, too. Specifically, I want to point out which ideas might be considered unusual, which might jar with the rest of pterosaur research, and highlight a few issues I’ve identified when combing the paper. I hope the following is taken in the spirit it’s intended — an honest response to a paper on an animal I’m deeply interested in — especially because two of the authors (Jim and John) are good friends. John is also aware of some of my misgivings so the following won’t come as a complete shock. In any case, I hope it’s clear that the intention here is not to whale on new research, but to highlight areas I predict will be contentious or did not find compelling, on the chance that they are of interest to others who find Quetzalcoatlus fascinating.
Old vs. new: Quetzalcoatlus 2021 compared to older reconstructions
My 2016 skeletal reconstruction of Q. lawsoni (then known as 'Q. sp') is now consigned to history. How did this now-six-year-old reconstruction hold up to the might of The Memoir? Read on...
One of the things I was keenest to see in Padian et al. (2021) was how my various reconstructions of Quetzalcoatlus have held up against new data. I’ve designed a lot of azhdarchids, including Quetzalcoatlus, for film and TV and wanted to see how close I’d landed to the reality of this animal using scraps of information gleaned from other papers. The artwork in Padian et al. (2021) is both copious and all excellent, stemming — of course — from the hand of John Conway, and we’re treated to a lot of skeletal reconstructions in multiple views. The paper largely focuses on Q. lawsoni, the smaller of the two named Quetzalcoatlus species, because virtually the entire skeleton of this animal is represented across multiple, similarly-sized specimens. As we've known since the 1970s, the charismatic giant wing that represents the giant Q. northropi can only take you so far in understanding Quetzalcoatlus: Q. lawsoni is really where the action is. And one thing to point out straight away is that our Q. lawsoni material is from several similarly-sized animals, but that there’s a lot of variation in limb metrics across them. They all seem to reach a 4.2-ish m wingspans through similar, but slightly different proportions (Andres and Langston 2021). This is interesting for all sorts of reasons, but also complicates any attempt at reconstruction. Probably the most obvious solution is to figure out what an ‘average’ Q. lawsoni looked like and work from that, although it would be neat to compare the extremes of proportion across the dataset too (something we're not doing here today).
The Padian et al. (2021) reconstruction of Q. lawsoni "posed in quadrupedal terrestrial stance". The half-crouched limbs are not artistic whimsy, but tied into ideas of Q. lawsoni hindlimb motion. We're going to get into that in a moment.
Comparing my old reconstructions with the new data, I think (if I may say so) that my work stands up relatively well. In honesty, I was surprised by Padian et al.’s (2021) assertion that: “there has never been a justification for the proportions of the bones used in any [Quetzalcoatlus] illustration”. This simply isn’t true because, despite the embargo over Quetzalcoatlus material, a lot of information on Q. lawsoni has been published over the last 50 years. These include mostly accurate limb metrics (Unwin et al. 2000); a full skull description (Kellner and Langston 1996) and dimensions of the cervical vertebrae (Steel et al. 2007; Witton and Naish 2008). There have even been pretty decent skeletal reconstructions based on examination of the original fossils (Paul 2002). These data are why the skeletal reconstruction published by Padian et al. (2021) isn’t massively different to some carefully researched pre-2021 versions. Compared to my own work, the only major discrepancies I found concern some posterior cervical lengths, the length of the body, and the size of the wing metacarpal. On the latter, my skeletal used a 620 mm length derived from Unwin et at al. (2000), which it now appears is too long: actual Q. lawsoni WMC lengths were in the range of 420-470 mm. Overall, Quetzalcoatlus was a little shorter in the arm than I’m used to, and fractionally longer in the neck and body, but it’s not a total visual transformation. Other distinctions between my older work and John’s new skeletal are just matters of opinion. For instance, the Padian et al. Quetzalcoatlus has a very tapered posterior skull, which I think is unlikely given the general condition of azhdarchoid crania. Specifically, completely known skulls from the azhdarchid Zhejiangopterus linhaiensis and at least one member of the azhdarchid sister clade, Chaoyangopteridae, have tall, sheet-like frontoparietal bones extending over and beyond their braincases (Cai and Wei 1994; Lü et al. 2008), and we see similar conditions in thalassodromids/ines as well. I thus regard this condition as likely for Q. lawsoni, but this will remain nothing more than opinion until we find a more complete skull.
Is the crouching pose of the Padian et al. Q. lawsoni reconstruction necessary? Ignoring the hindlimb restoration philosophy (see below), there may be scaling issues with the forelimb affecting things too: an 'adjusted' skeletal to the right shows that Q. lawsoni could stand tall without issue.
But in checking out the new reconstructions I also noted some less subjective differences. One of the more striking aspects of Quetzalcoatlus 2021 is the proposed habitual crouching pose. It reflects both assumptions about the hindlimb articulations (which we’ll discuss at length below) as well also the unexpected shortness of the forelimb. But even accounting for that short wing metacarpal, the wing looked strangely stunted to me. Upon investigation, I found that the wing skeleton is probably incorrectly scaled. Specifically, when compared to metrics given in Padian et al. (2021) and Andres and Langston (2021), the reconstructed radius/ulna and wing metacarpal lengths are 11 and 10% shorter (respectively) than an ‘average’ Q. lawsoni wing. This makes the arm quite a lot shorter than it should be and, when adjusted, there’s no problem making Quetzalcoatlus stand in a more typical, fully-upright posture. Indeed, the forelimb becomes long enough that the entire hindlimb can be extended vertically under the body without the arm looking over-extended (above).
The size of the foot also drew my attention. It’s been remarked that azhdarchids had small feet (Cai and Wei 1994; Hwang et al. 2002; Witton and Naish 2008; Andres and Langston 2021) and yet Padian et al. (2021) show Quetzalcoatlus as a relatively large-footed animal. The diminutive foot size of azhdarchids was one reason Darren Naish and I suggested they were terrestrial foragers back in 2008 (Witton and Naish 2008) and when I saw the big, flappy feet of 2021's Q. lawsoni I thought we’d got things wrong. But, again, there’s a measuring complication here. Padian et al. (2021) suggest the metatarsus (the long bones of the foot) was about 150 mm long, which is about 25% of the tibiotarsus length, and this is what's shown in the reconstruction. But Andres and Langston (2021) give a very different set of figures: they record the metatarsus as 15% of the tibiotarsus and 82.5-90 mm long. Andres and Langston further stress that Quetzalcoatlus had the third-shortest foot, relative to body size, of any known pterosaur, and this emphasis makes me think their measurements are more likely to be correct. If so, and we then assume — as suggested by Padian et al. — that the toes were a similar length to the metatarsals, Q. lawsoni would have had tiny feet of c. 160-180 mm long. This is a little over half of what's reconstructed for the Padian et al. restoration, but similar to the foot proportions of Zhejiangopterus.
My 2022 skeletal reconstruction of Quetzalcoatlus lawsoni, incorporating the adjusted proportions outlined above.
Once Quetzalcoatlus 2021 is adjusted to suit these adjusted measurements, it looks a lot less strange. You can get a sense of this from my own rebuilt skeletal reconstruction of Q. lawsoni, above. It should be stressed that there is a defence to these scaling issues: the aforementioned variation in limb metrics where specimen proportions can vary by over 10%. So perhaps Padian et al. haven’t reconstructed an ‘average’ Q. lawsoni, but they’ve still reconstructed something within the proportions of this species? There may be some validity to this, but comparing the presented reconstruction to the metrics of Andres and Langston (2021) suggests it's still something of a stretch. Nevertheless, the weird variation in Q. lawsoni proportions may be where all these issues originated.
Deja Qu, part I: crouching Quetz, hidden controversy
Let’s now look beyond proportions to functional morphology, starting with that strange crouching hindlimb. This reflects the idea that the Quetzalcoatlus femur was perpetually held subhorizontally with a maximum downward rotation of only 70-75𝆩 (Padian et al. 2021). This, it's said, prohibits the femur from swinging backwards under pelvis as is widely interpreted and illustrated for pterosaurs across scientific literature and palaeoartworks, and it's not a new idea: it's taken straight from Padian papers published in the 1980s. The arguments are principally the same: that the articular surfaces of the pterosaur knee do not allow the leg to straighten, and that the femoral curvature of pterosaurs recalls that of birds, implying a subhorizontal orientation (see Padian 1983a, b; Padian et al. 2021). Padian et al. (2021) also mention that their proposed posture scores points for fitting Jurassic pterosaur tracks from Crayssac, France, an idea that also ties into classic Padian literature. Why fit Quetzalcoatlus into the tracks of relatively tiny Jurassic pterodactyloids and not the Haenamichnus tracks widely considered to have been made by a Korean azhdarchid (Hwang et al. 2002)? Because Kevin Padian (2003, 2008; Padian and Olsen 1984) has long been sceptical about the origins of pterosaur tracks, cumulating in the belief that only examples found in southern France are genuine pterosaur ichnites. Most or all others, he argues, were left by other reptiles; chiefly, crocodylian-like ones.
Suggested ranges of motion at the Q. lawsoni hip and knee, according to Padian et al. (2021). The precision drawings and figures suggest a lot of confidence in these data, but they contrast with several comments about the poor quality of the Q. lawsoni pelvis in the memoir, and the difficulty of reconstructing it accurately. The proposed range of knee articulation is also very restricted compared to analyses of this joint in other pterosaurs.
Understanding that Padian et al. (2021) has been written from this perspective explains why its discussion of hindlimb mechanics frequently jars against more recent studies. Padian et al. concede that the concept of subhorizontal pterosaurian femora contrasts with the conclusions of at least one team (Costa et al. 2014), but don't mention the heaps of other investigations it also conflicts with (e.g. Bennett 1990, 1997, 2001; Unwin 1996; Fastnacht 2005; Wilkinson 2008). While it would be incorrect to say that we understand everything about the motion of the pterosaur hindlimb, most researchers are pretty happy that the femur could swing into a subvertical pose. Indeed, some studies conclude that this is the optimal position for the pterosaur hindlimb when walking, providing the best mechanical advantage for the muscles that swing the leg forward and back (Fastnacht 2005; Costa et al. 2014). It’s also generally observed that the articular surfaces of pterosaur knees extend to the tips of the limb bones, allowing them to adopt almost entirely straightened knee poses (e.g. Bennett 2001; Wilkinson 2008). Padian et al. (2021) provide the first assessment of this for azhdarchid knees, but I admit to wondering why they think the condyles are so limiting when azhdarchid hindlimb joints look pretty similar to those of other pterodactyloids (see Godfrey and Currie 2005; Averianov 2010; Andres and Langson 2021).
With the weight of opinion being that pterosaur hindlimbs were actually pretty different to bird legs, I'm surprised the Q. lawsoni functional analysis leads so strongly with its assessment of a bird-like subhorizontal femur. At very least, those other studies warrant discussion. And as for the seeming validation that a crouching Quetzalcoatlus can be made to fit Jurassic pterosaur trackways, this is a moot point: conventional, upright hindlimb postures fit these tracks too (Bennett 1997; Mazin et al. 2003). The take-home here is that the proposal of Quetzalcoatlus having a subhorizontal femur, and thus being limited to a strange, crouching pose, is both odd and not well substantiated against the consensus view of pterosaur research. It really needs bolstering with more data to be credulous.
Deja Qu, part II: Leg folding…
I mentioned above that some disagreement exists among the Padian et al. (2021) team on several topics, two of which concern flight pose and membrane shape. While Jim and John advocate something approximating the classic sprawled-leg flight pose and at least some degree of hindlimb membrane attachment, Padian prefers a bird-like configuration where the hindlimb is tucked underneath the body and the wing membrane anchors at the hip. Again, these latter ideas are Padian hypotheses that first aired 40 years ago (e.g. Padian 1983b; 1988). Predicting pterosaur membrane shapes remains a complex issue and is beyond our scope for discussion here: it’ll suffice to say that there is no evidence for a pelvic membrane attachment in any pterosaur, and that the handful of inboard membrane fossils we have collectively point to a distal hindlimb attachment across Pterosauria, including in Azhdarchoidea. This was well documented by Ross Elgin et al. (2011), a paper which Padian et al. cite and (probably unfairly) dismiss with just a few words. As for the question of flight pose: this boils down to whether Quetzalcoatlus was incapable of adopting the classic ‘sprawled-leg’ posture widely reconstructed for pterosaurs and, if not, did it have to adopt an unusual, avian-like one instead?
Q. lawsoni wing poses illustrated by Padian et al. (2021): which do you prefer? John Conway and Jim Cunningham are on record preferring model C, while Kevin Padian argues for D. My vote, given what fossils show of pterosaur wing membranes, would be for something between B and C (distal hindlimb membrane anchor, but a tighter trailing curve than B).
As alluded to above, discussions over pterosaur pelvis-hindlimb arthrology are nothing new. How far pterosaurs could move their femora around has been the subject of a large number of papers (e.g. Padian 1983a, b, 2003; Wellnhofer 1988; Bennett 1990, 1997, 2001; Unwin 1996; Wilkinson 2008; Costa et al. 2014; Frigot 2018) leading to a general consensus that most or all pterosaurs could move their hindlimb through a wide range of motion, walking and standing with a near-vertical femur but also swinging their legs out in flight. This conclusion is not just based on manually articulating bones but also on hundreds of articulated fossils showing pterosaurs preserved with both upright and splayed hindlimbs. These include two Zhejiangopterus specimens with butterflied hindlimbs (illustrated in Cai and Wei 1994 and Witton 2013) that show azhdarchids conforming to pterosaur norms. If Quetzalcoatlus was incapable of adopting a hindlimbs-out flight pose, it would have been highly aberrant and we’d need good evidence of such an interpretation: ideally, a well-preserved pelvis with an uncrushed acetabulum (hip socket) and a correspondingly well-preserved femur that allowed us to demonstrate, beyond doubt, limited capacity for hindlimb abduction.
The Q. lawsoni pelvis as illustrated by Andres and Langston (2021): it's far from the best-preserved piece of our Q. lawsoni inventory. Are these the sort of pelvic remains we can use to substantiate a radical departure from our typical interpretations of pterosaur hindlimb arthrology? Probably not.
But, alas, here’s how Andres and Langston (2021) describe the only recovered pelvic material of Q. lawsoni: “fractured, heavily encrusted with concretionary material, and [with] matrix… often stained a similar color to the bone.” They conclude that “this pelvic plate is not preserved well enough to decisively determine its orientation with respect to the vertebral column” and that the angle of the acetabulum cannot be interpreted with confidence. Even Padian et al. (2021) concede that “the pelvis cannot be reconstructed in three dimensions with confidence”. There's an agreement, then, that we can't put the Q. lawsoni pelvis together with certainty, and this is a problem. We've learned from multiple studies that restoring pterosaur leg mobility is influenced by numerous factors including the shape and orientation of the hip socket, the precise angle of the pelvis with respect to the spinal column, the articulation of the pelvic bones themselves, and the inclination of the torso (e.g. Wellnhofer 1988; Bennett 1990; Fastnacht 2005; Wilkinson 2008; Costa et al. 2014). In other words, we need really, really excellent fossils to even start thinking about such investigations and if we can't reconstruct the Q. lawsoni pelvis, we cannot say much about the range of motion of the leg.
It's for this reason that Padian et al. can only infer avian-like hindlimb mobility for Q. lawsoni, which they openly declare in their introduction to this topic: “Given the bird-like features of the entire hind limb, which not only bear anatomical resemblance but speak to functional similarity, it appears reasonable to begin with the kinds of postures and degrees of movements found in birds”. This is surely not the right approach and certainly undermines the suggestion that "In flight, it is most plausible that the hind limbs were drawn up bird-like, with the knee anterior to the acetabulum". Alternatively, if we can't model the hindlimb arthrology for Quetzalcoatlus, we have to fall back on what we're learned from other pterosaur species, not point to an anatomically distinct, phylogenetically distant pterosaur relative and made sweeping inferences. In all, I find nothing compelling about the concept of Quetzalcoatlus having to tuck its legs up like a bird, and I 100% agree with Jim and John in their endorsement of a more traditional, hindlimb-splayed flight configuration.
…and wing folding
Discussing flight brings us to another potentially contentious topic: the Q. lawsoni wing, or, rather, wing folding. We know a fair bit about how pterosaurs collapsed their wings for standing and walking (e.g. Wellnhofer 1988; Unwin 1996; Bennett 1997, 2001; Wilkinson 2008) and the general conclusion is that pterosaur forelimb articulation was complex. Their arm joints didn’t articulate uniaxially (i.e. in one plane); instead, the elbow and wrist deflected their distal limb bones medially and laterally as they opened and closed. We've found that, to get pterosaurs walking in their trackways, the pterosaur elbow needed to stick out from the body a little and that (like many dinosaurs) the palms of pterodactyloid hands faced inwards, as if the hands were ready to clap for a round of applause*. This is why pterodactyloid trackways show handprints with sideways projecting fingers: the digits have swung under the big knuckle of the wing digit to extend away from the body. The wing finger itself follows the same rules, so it folds up along the outside of the wing. A quirk of the wing metacarpal joint means that the wing digit is somewhat posteriorly deflected when it does this, stowing alongside the forearm during terrestrial progression. These basic findings are something that we’ve modelled from pterodactyloid bones and also witnessed in dozens, maybe hundreds, of well-preserved pterosaur fossils. Whenever we have an articulated, tightly folded pterosaur wing, the wing finger lies over the radius and ulna, not under it, and the palm of the hand faces inwards. We know this applies to azhdarchids too, thanks to articulated fossils of Zhejiangopterus (Cai and Wei 1994). We also have azhdarchid tracks, Haenamichnus, showing their hands were orientated in a typical, ‘palms inward’ pterosaur fashion when walking (Hwang et al. 2002). All expectations are, therefore, that Quetzalcoatlus would follow this familiar configuration.
*The situation is different in non-pterodactyloids, but that’s another story.
One of my favourite images for showing the complexity of pterosaur forelimb articulation, from Wilkinson (2008). The pose here can be regarded as 'extreme' as we have good data indicating that pterosaurs stood more upright than this, but the orthographic views show how the forearm and hand are deflected as the wing folds. Note how the wing finger and walking fingers rotate around an axis parallel to the midline of the body, allowing the fingers to project sideways while the wing finger folds against the outer arm.
Given these relatively well-established models, it’s something of a surprise to see Padian et al. (2021) showing Q. lawsoni doing something different. The wing is positioned so that the palm of the hand faces somewhat forward, allowing the wing digit to tuck under the elbow, despite the walking fingers still projecting laterally. The ability to draw the wing finger under the elbow is, apparently, a consequence of a slight downward deflection to the end of the wing metacarpal which changes the orientation of the joint, but I suspect it was also influenced by the methodology for modelling the standing pose. It’s reported that this was deduced by manually positioning casts of Q. lawsoni fossils in a plausible upright arrangement, an exercise which "began by placing the distal end of the wing metacarpal... with its distal condyles oriented posteriorly (so that the wing finger could be directed behind the elbow and close to the body wall)". It seems it was decided, a priori, that this is where the wing finger should go.
Proposed wing folding of Q. lawsoni, from Padian et al. (2021).
It’s a shame that no photos or diagrams of this work were published because, while those of us who have not handled the Quetzalcoatlus bones can’t really say that this interpretation is wrong, there are lots of legitimate questions about it that make me hesitant in accepting it outright. For example, Quetzalcoatlus is not unique for having that slight downturn at the end of the wing metacarpal: we see similar conditions in taxa like Pteranodon and Tapejara. In Pteranodon at least, they’ve been factored into arrangements of the folded pterosaur forelimb and do not result in the wing finger tucking under the elbow (Bennett 2001). I'm also not clear how the Q. lawsoni walking fingers are depicted as splaying out to the side when their respective metacarpals are positioned on the front of the wing: unless these joints were strangely bevelled, surely they should be facing more posteriorly? It's strange to have the palm facing forward anyway, as studies exploring the impact of angling the pterosaur palm forward find that such poses are only possible if the forelimb adopted a crazy, implausible configuration (Bennett 2001).
Exploring how pterosaurs stood has taken us down some strange roads. In 2001, Chris Bennett attempted to pose Pteranodon in a once traditional configuration with a forward-facing hand: it didn't go well (note that the walking fingers are even upside down!).
And there are other issues, too. I wonder why the wing casts exercise was reconstructed with the wing finger joint facing posteriorly rather than, as we'd expect, laterally. In such a configuration the wing finger is almost certainly going to tuck under the elbow, because the elbow has to bow out from the shoulder when a pterosaur stands. I think that would happen regardless of pterosaur taxon. Also of relevance here is that the Q. lawsoni wing metacarpals are, reportedly, poorly-preserved at the proximal (wrist) ends: as with our discussion of the hip bones, above, I wonder if the material is well-enough preserved to substantiate such bold claims? And what of Zhejiangopterus, with its well-behaved wing finger? Why, again, is Q. lawsoni so different to other azhdarchoids?
Zhejiangopterus linhaiensis as illustrated by Cai and Wei (1994). OK, this is hardly the height of palaeontological visualisation (to be fair, the original fossil is barely more than an outline) but you can see lots of important functional features in this articulated azhdarchid specimen including splayed hindlimbs, medially-facing palms, and wing fingers that fold up against the outside of the wing. This is all good data that any interpretation of Q. lawsoni functionality needs to be considered against: it's our only direct insight into how azhdarchid skeletons fitted together.
In sum, I'm not saying that an elbow tucked interpretation is outright wrong. I am, however, very sceptical given the above points and would want to see further research — ideally informed by previous studies on pterosaur wing folding, and bringing in data from Zhejiangopterus and Haenamichnus as well— to substantiate an elbow-tucked wing finger. As you'll note in the art above, I've stuck to convention on this matter with my latest Q. lawsoni piece.
Bipedal launching: back on the table?
Finally, another area of contention between the authors of Padian et al. concerns launch strategy: how did Quetzalcoatlus become airborne? Here, the split is once again between Jim and John on one side, who advocate quadrupedal launch, and Kevin Padian on the other, who prefers a bipedal launch model. This split is not surprising because, some years ago, Jim independently drew the same conclusion about pterosaur launch as Mike Habib, who wrote the first paper on flying reptile quad-launch in 2008. Since then, this idea has become the pterosaur launch hypothesis to beat. As outlined at length in this post, it’s the only concept that explains (and with substantial quantification and experimentation) everything we understand about pterosaur size, proportions and muscle volumes, while also fitting launch expectations from pterosaur trackways (i.e. that, among living animals, the gait used for terrestrial locomotion is the same gait used for take-off). It also avoids having to downsize pterosaurs to ridiculously small masses to achieve flight, as exemplified by Chatterjee and Templin’s (2004) conclusion that a giraffe-sized azhdarchid must mass 75 kg or less to facilitate take-off. Under quad-launch theory, giant pterosaurs can easily be 200 or 300 kg and still become airborne (Habib 2008, 2013; Witton and Habib 2010; Habib and Cunningham 2013).
A Q. lawsoni standing-start bipedal launch, illustrated in Padian et al. (2021). Is this a viable launch mechanic for a 4.2 m wingspan pterosaur? Opinion is split among the Padian et al. authorship.
It’s against this that Padian et al. refloat the idea of Quetzalcoatlus being a bipedal launcher, an idea also emphasised in press releases. The discussions of both bipedal and quadrupedal launch in Padian et al. (2021) are qualitative, mostly focusing on how the skeleton of Quetzalcoatlus can be moved into various launching postures, and there’s little engagement with what’s been said about pterosaur launch by recent workers. For example, Padian et al. suggest that Quetzalcoatlus might have lacked forelimb bending strength to sustain quad launch (i.e. that the forelimb would fail under such stress), ignoring the fact that Mike Habib and I demonstrated over a decade ago that the Q. lawsoni humerus was five times stronger under bending than the femur, and that even the neck bones of Quetzalcoatlus were stronger than its legs (Witton and Habib 2010). There is no explanation for why Quetzalcoatlus lacks the robust hindlimb anatomy of a large hindlimb launcher, when Mike has demonstrated that any flying animal above 500 g starts augmenting its launch limb anatomy to achieve sufficient power and reinforcement to sustain take-off (Habib 2008). And there’s no discussion of why previous calculations of bipedal launch, which have been universally hamstrung by having to lower pterosaur masses to ridiculous levels, went wrong. Padian et al. prefer a (relatively low) mass of 150 kg for a giant azhdarchid, but even this would be two- or three-times too heavy for any published bipedal launch mechanic. The fact bipedal launch proponents have consistently failed to get realistically-massed pterosaurs airborne isn't something we can just ignore: it's evidence against this hypothesis. There’s more we could say, but you're getting get the idea: this attempted resurrection of bipedal launch as a viable take-off mechanic for even the small Quetzalcoatlus species has not, in my view, been well-argued, and does nothing to displace quad-launch as the superior pterosaur take-off hypothesis.
Conclusion: extraordinary claims... etc., etc...
And that, I think, is all I want to say on this for now. In short, I can't buy that Quetzalcoatlus is anywhere near as strange as the conclusions of Padian et al. (2021) imply. And to be clear, there is nothing wrong with arguing that Quetzalcoatlus was an aberrant species, but if you're going to make such assertions you need to present excellent, thorough and fully watertight analyses, and I just don't think Padian et al. do this. There are too many unaddressed complications, overlooked counterarguments and obvious questions raised around their more unusual hypotheses to take them as read, and the fact that a lot of the ideas are somewhat forgotten, decades-old hypotheses is something that really needed addressing: if we've moved on from those ideas once, why are they suddenly viable now? What's changed to make hip-anchored membranes and bird-like knees plausible for pterosaurs in the 2020s? It's this lack of consideration and engagement with modern pterosaur science that is at the core of my scepticism with so much of the paper. But this, of course, is only my take: it’ll be interesting to see what other researchers make of this now that Quetzalcoatlus is finally, and happily, available for unrestricted research access.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Andres, B., & Langston Jr, W. (2021). Morphology and taxonomy of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41(sup1), 46-202.
Averianov, A. O. (2010). The osteology of Azhdarcho lancicollis Nessov, 1984 (Pterosauria, Azhdarchidae) from the late Cretaceous of Uzbekistan. Proceedings of the Zoological Institute RAS, 314(3), 264-317.
Bennett, S. C. (1990). A pterodactyloid pterosaur pelvis from the Santana Formation of Brazil: implications for terrestrial locomotion. Journal of Vertebrate Paleontology, 10(1), 80-85.
Bennett, S. C. (1997). Terrestrial locomotion of pterosaurs: a reconstruction based on Pteraichnus trackways. Journal of Vertebrate Paleontology, 17(1), 104-113.
Bennett, S. C. (2001). The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon Part II. Size and functional morphology. Palaeontographica Abteilung A, 113-153.
Cai, Z & Wei, F. (1994). On a new pterosaur (Zhejiangopterus linhaiensis gen. et sp. nov.) from Upper Cretaceous in Linhai, Zhejiang, China. Vertebrata PalAsiatica, 32(03), 181.
Chatterjee, S., & Templin, R. J. (2004). Posture, locomotion, and paleoecology of pterosaurs (Vol. 376). Geological Society of America.
Costa, F. R., Rocha-Barbosa, O., & Kellner, A. W. A. (2014). A biomechanical approach on the optimal stance of Anhanguera piscator (Pterodactyloidea) and its implications for pterosaur gait on land. Historical Biology, 26(5), 582-590.
Elgin, R. A., Hone, D. W., & Frey, E. (2011). The extent of the pterosaur flight membrane. Acta Palaeontologica Polonica, 56(1), 99-111.
Fastnacht, M. (2005). The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones. Acta Palaeontologica Polonica, 50(2).
Frigot, R. A. (2018). Pelvic musculature of Vectidraco daisymorrisae and consequences for pterosaur locomotion. Geological Society, London, Special Publications, 455(1), 45-55.
Godfrey, S.J., & Currie, P.J. (2005). Pterosaurs. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis, p. 292-311.
Habib, M. B. (2008). Comparative evidence for quadrupedal launch in pterosaurs. Zitteliana, B28, 159-166.
Habib, M. (2013). Constraining the air giants: limits on size in flying animals as an example of constraint-based biomechanical theories of form. Biological Theory, 8(3), 245-252.
Habib, M. B., & Cunningham, J. (2010). Capacity for water launch in Anhanguera and Quetzalcoatlus. Acta Geoscientica Sinica, 31, 24-25.
Kellner, A. W., & Langston Jr, W. (1996). Cranial remains of Quetzalcoatlus (pterosauria, Azhdarchidae) from late cretaceous sediments of big bend national park, Texas. Journal of Vertebrate Paleontology, 16(2), 222-231.
Hwang, K. G., Huh, M. I. N., Lockley, M. G., Unwin, D. M., & Wright, J. L. (2002). New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri formation, southwestern Korea. Geological Magazine, 139(4), 421-435.
Lü, J., Unwin, D. M., Xu, L., & Zhang, X. (2008). A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution. Naturwissenschaften, 95(9), 891-897.
Mazin, J. M., Billon-Bruyat, J. P., Hantzpergue, P., & Lafaurie, G. (2003). Ichnological evidence for quadrupedal locomotion in pterodactyloid pterosaurs: trackways from the Late Jurassic of Crayssac (southwestern France). Geological Society, London, Special Publications, 217(1), 283-296.
Naish, D., & Witton, M. P. (2017). Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators. PeerJ, 5, e2908.
Padian, K. (1983a). Osteology and functional morphology of Dimorphodon macronyx (Buckland) (Pterosauria: Rhamphorhynchoidea) based on new material in the Yale Peabody Museum. Postilla, 189, 1-44.
Padian, K. (1983b). A functional analysis of flying and walking in pterosaurs. Paleobiology, 9(3), 218-239.
Padian, K. (1988). The flight of pterosaurs. Natural History, 12, 58-65.
Padian, K. (2003). Pterosaur stance and gait and the interpretation of trackways. Ichnos, 10(2-4), 115-126.
Padian, K. (2008). Were pterosaur ancestors bipedal or quadrupedal?: morphometric, functional, and phylogenetic considerations. Zitteliana, B28, 21-33.
Padian, K., Cunningham, J. R., Langston Jr, W., & Conway, J. (2021). Functional morphology of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology, 41(sup1), 218-251.
Padian, K., & Olsen, P. E. (1984). The fossil trackway Pteraichnus: not pterosaurian, but crocodilian. Journal of Paleontology, 178-184.
Paul, G. S. (2002). Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds. JHU Press.
Steel, L., Martill, D. M., Kirk, J. R. J., Anders, A., Loveridge, R. F., Frey, E., & Martin, J. G. (1997). Arambourgiania philadelphiae: giant wings in small halls. The Geological Curator, 6, 305-313.
Unwin, D. M. (1996). Pterosaur tracks and the terrestrial ability of pterosaurs. Lethaia, 29(4), 373-386.
Unwin, D. M., Lü, J., & Bakhurina, N. N. (2000). On the systematic and stratigraphic significance of pterosaurs from the Lower Cretaceous Yixian Formation (Jehol Group) of Liaoning, China. Fossil Record, 3(1), 181-206.
Wellnhofer, P. (1988). Terrestrial locomotion in pterosaurs. Historical biology, 1(1), 3-16.
Wilkinson, M. T. (2008). Three-dimensional geometry of a pterosaur wing skeleton, and its implications for aerial and terrestrial locomotion. Zoological Journal of the Linnean Society, 154(1), 27-69.
Witton, M. P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press.
Witton, M. P., & Habib, M. B. (2010). On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on pterosaur flightlessness. PloS one, 5(11), e13982.
Witton, M. P., & Naish, D. (2008). A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS one, 3(5), e2271.
Witton, M. P., & Naish, D. (2013). Azhdarchid pterosaurs: water-trawling pelican mimics or “terrestrial stalkers”?. Acta Palaeontologica Polonica, 60(3), 651-660.
Tyrannosaurus engages in some closed-mouth vocalising, but which species of Tyrannosaurus is this? It might seem that, up until recently, there would be only one answer, but this is actually an old question dating back to at least the early 1970s. Hey, writing about this sounds like a fun idea for a blog post. Oh look, someone else thought so too - read on...
Even aliens living on the far side of the Moon are aware that, last month, independent researcher and palaeoartist Gregory S. Paul led a team of authors proposing that our traditional take on Tyrannosaurus rex was wrong. According to Paul et al. (2022), this classic genus does not contain just one species, but actually three: the stocky, geologically older T. imperator, and two descendant, coexisting younger species, T. rex and T. regina. As documented in the New York Times and elsewherein numerousarticles, the response from tyrant dinosaur experts was not enthusiastic; to the contrary, most workers concluded that the study was underwhelming and unlikely to change the status quo. It’s not my intention to provide a detailed breakdown on this paper as relevant experts have already done this in recent weeks on various news and social media sites. It'll suffice to say that, while some of the ideas floated by Paul et al. are certainly interesting, the vague, very short diagnoses of the three species, the failure to provide strong statistical support for “robust” and “gracile” morphotypes within our T. rex inventory, and the inability to assign several well-preserved partial skeletons to the new species will likely prove sufficiently problematic to prevent widespread adoption of this new scheme. I hear a response is already being written.
But one, perhaps surprising, reaction to Paul and colleagues’ (2022) research is that most experts really weren’t shocked by the idea of several species within the T. rex hypodigm, and several even remarked that the idea itself is plausible even if our data, for the time being, do not support it. While the exact geological longevity of Tyrannosaurus rex is open to question (Paul et al. 2022), it was at least around for close to one million years (Brusatte and Carr 2016) and existed in varying habitats across a wide spread of western North America, from southern Alberta to the Texas/Mexico border, and from Utah to the Dakotas (Sampson and Loewen 2005). Compared to other theropods, that’s a broad spatiotemporal spread and one that could conceivably contain hitherto unappreciated variation that requires close analysis to uncover.
And it’s this topic of looking closely at our T. rex sample that I want to discuss today. It might seem that Paul and colleagues’ proposal is a radical dismantling of a fossil icon, and, indeed, this status is emphasised by Paul et al. themselves, who remark in their supplementary data and press work that they are “likely to displease many who are enamoured with the tyrant lizard king Latin moniker”, but that their "mini-revolution" will “force people to face the issue”. But, viewed more broadly, their study is actually only the latest in a long line of research that has expanded or shrunk our inventory of T. rex fossils and toyed with the idea of multiple Tyrannosaurus species. Indeed, even the features said to characterise the T. regina, rex and imperator are familiar from older publications (e.g. Larson 2008a). The result is that, when reviewing older literature on Tyrannosaurus taxonomy in light of these recent discussions, it’s difficult to avoid a sense that these seemingly-new ideas are not only re-treading old ground, but maybe even tripping over it.
Graphic summary of the classification of Tyrannosaurus specimens (cranial material alone shown here) in the late 20th century. This illustration and those that follow do not represent the entirety of T. rex material known at any one time, but focus on named T. rex specimens that have been proposed as distinct taxa at one time or another.
Despite the long-held celebrity of Tyrannosaurus and it being known to science for over a century, most of what we’ve learned about this animal has been discovered in the last 30 years. In fact, little of note was published on T. rex during the mid-20th century and only a few significant specimens were uncovered (Larson 2008b). This corresponds with little taxonomic work and means we can skip ahead to start this story around 1970. Tyrannosaurus fans will realise that this omits discussion of the historic taxa “Dynamosaurus” and “Manospondylus” but, given that they were satisfactorily dismissed by Osborn (1906, 1917) and have no chance of being revived today, we can gloss over the earliest history of T. rex studies with only one important note: rushed as it was, Henry Osborn’s (1905) naming and type designation for Tyrannosaurus rex were extremely solid. So remember that, in spite of everything that follows in this article, and whatever else future generations have to say about the number of Tyrannosaurus species, T. rex itself is not going anywhere.
Rushing through that earliest period of Tyrannosaurus research puts us in the mid-20th century when, following a world war-induced hiatus in North American palaeontology, collectors and researchers returned to the field during the 1960s and ‘70s to resume exploration of American fossil resources. Among many important discoveries were new specimens of Tyrannosaurus and, at this point in history, each one was significant because T. rex was still only represented by a few examples. As our sample size grew from a few to a handful, and then to a dozen and more, it became apparent that no two T. rex individuals were exactly alike: T. rex evidently held a large amount of intraspecific variation within its population (Carpenter 1990). With this observation came the question of how much difference could be permitted in these fossils before specimens could no longer be identified as T. rex. The first rumblings on this never made it to print but were included in an unpublished 1972 thesis by then-student Douglas “Quetzalcoatlus” Lawson, who named Tyrannosaurus“vannus” for a relatively small, seemingly unusual tyrannosaurid maxilla from the Texan Javelina Formation. T. “vannus” never became “official” in the eyes of zoological nomenclature because it was never formally published and, in any case, Lawson revised his interpretation soon after, referring the same bone to T. rex itself (Lawson 1976). But in a later, 1990 review of T. rex variation, Kenneth Carpenter independently opined that this maxilla might represent a distinct southern Tyrannosaurus species. Others (Brochu 2003; Carr 2020) have been less convinced that it represents a new species, with Brochu (2003) suggesting that if the Texan maxilla was not T. rex, it must still represent a very close relative.
FMNH PR2081, better known as "Sue", represents one extreme of Tyrannosaurus form in being an especially large and robust example of the genus. These features have seen this specimen (shown here in its older, more prominent place in the Field Museum of Natural History) at the centre of many discussions of T. rex diversity and disparity. Photo by Connie Ma, from Wikimedia, CC BY-SA 2.0.
This was only the start of the buzz circulating around the concept of Tyrannosaurus having multiple species, which gained further traction in the 1980s. Again, the amount of variation in T. rex specimens was generating conversation, and both Horner and Lessom (1993) and Larson (2008) give Robert Bakker credit for observing features in Tyrannosaurus skeletons that might distinguish certain morphotypes from others. It’s reported that Bakker was so confident in this that only a slightly increased T. rex sample size might have seen him formally propose more Tyrannosaurus species before the turn of the millennium. It was perhaps these rumours and conversations that prompted Paul (1988) to muse on multiple Tyrannosaurus species in his influential book Predatory Dinosaurs of the World. Despite acknowledging variation in T. rex dentition and limb structure, Paul concluded that no, the Tyrannosaurus specimens known in the late 1980s were not diverse enough to represent more than one species. But other researchers weren’t so sure. Molnar (1991) responded to Paul’s comments by questioning whether we had enough T. rex specimens to draw any firm conclusions about the number of species they really represented, and suggested that the question would remain open until we acquired more data. Others were more confident that Paul was wrong. Donald Glut (1997) recorded that researcher Stephen Pickering privately issued manuscripts in 1996 proposing that several T. rex specimens, including the famous “Sue” skeleton, represented a new species that he named Tyrannosaurus "stanwinstonorous" after the special effects pioneer responsible for the animatronic dinosaurs in Jurassic Park and its sequels. Pickering argued that multiple features of the skull and the trait of adults being 6-7 % larger than Tarbosaurus or T. rex distinguished "stanwinstonorous" from other tyrannosaurs, but — as with Lawson’s T. “vannus” — lack of widespread publication prevented this name from entering scientific consideration.
So, even as palaeontologists circled Tyrannosaurus looking for specimens to split from the T. rex hypodigm in the late 20th century, no formal proposals were published. But taxonomic acts relevant to T. rex were being carried out as smaller, often fragmentary North American tyrannosaurs that we’d eventually recognise as Tyrannosaurus were receiving names or reclassification. With some of these specimens having been found decades prior, fairly convoluted taxonomic histories are associated with them (see Carr and Williamson 2004 for a summary). Here, let’s just cut to the chase and list them with their final, accepted binomials: Dinotyrannus megagracilis (Paul 1988; Olshevsky and Ford 1995), Stygivenator molnari (Paul 1988; Olshevsky and Ford 1995) and — most famous of all — Nanotyrannus lancensis (Gilmore 1946; Bakker et al. 1988). Despite recognition that at least some of these species (D. megagracilis and S. molnari) were based on juvenile or subadult material, the idea that they may represent subadults of T. rex was not considered, and the validity of N. lancensis was hinged, to some extent, on it being a subadult or adult that reached maturity at a smaller size and lighter frame than T. rex itself (Paul 1988; Bakker et al. 1988). This thinking reflects, in part, a period in vertebrate palaeontology dominated by typological approaches where specimens were assigned to species based on morphology alone without factoring in the influences of growth or individual variation. Under such schemes, it was rare, though not unheard of (Rozhdestvensky 1965; Russel 1970), to ally juvenile theropod specimens with adult specimens.
T. rex taxonomy made simple: everything is T. rex! There are, of course, dozens more specimens we could show here: this graphic only shows the skulls of specimens thought to represent different Tyrannosaurus species at one time or another.
A more holistic philosophy toward the classification of fossil species has, in the last few decades, replaced such thinking and allows us to realise that, for example, a young juvenile specimen might look very different to an old adult. This can make characterising fossil species more difficult, but the results are more rewarding as we get more detailed and realistic insights into the life histories and ecologies of ancient animals. Such practices have typified most recent taxonomic work on Tyrannosaurus, emphasising the need to investigate features of specimen growth stage (e.g. bone fusions, histological evidence of specific age etc.) alongside morphology when considering what is, and what isn’t, Tyrannosaurus rex (e.g. Carr 1999, 2020; Carr and Williamson 2004; Woodward et al. 2020).
These works have shown that Tyrannosaurus was even more variable than initially realised, not only being differentiated in build and size as an adult but also looking entirely different as a juvenile. As is now well-known to Tyrannophiles, T. rex grew from a dog-sized, slender-snouted hatchling to a supersized, gnarly-skulled adult with unprecedented robustness among theropods. Carr (2020) characterises T. rex growth as undergoing “secondary metamorphosis” at the onset of sexual maturity, in which it transformed from a slender, “bird-meets-horse” juvenile body plan to a robust, deep-snouted, massive-bodied adult form. The result is an interpretation of Tyrannosaurus rex that occupied a range of disparate anatomies throughout its lifespan, within which small tyrants like “D. megagracilis” and “S. molnari” can be absorbed into a well-substantiated growth sequence (Carr and Williamson 2004). And yes, while a minority favour Nanotyrannus as a distinct taxon (e.g. Larson 2013), it also plots neatly into this system without issue (Carr 1999, 2020; Currie 2003; Woodward et al. 2020).
It's strange to think that this fleet-footed predator is the same species as that 6-10 tonne tank we were recently discussed as a head-butter, but the evidence that T. rex underwent this radical transformation is very strong. This dramatic transformation is also surely one of the major complicating factors in our attempts to unravel Tyrannosaurus taxonomy.
This hypothesis does more than just tidy up T. rex taxonomy. It also establishes that Tyrannosaurus was probably not a “normal” theropod in terms of its ecological role, and that there may be a good reason why this one species occupied such an expanse of time and space in Maastrichtian North America. The 30-odd year lifespan of one Tyrannosaurus encapsulated the ecological potential of several grades of predatory dinosaurs (Holtz 2021), and we might expect such an adaptable animal to have a long evolutionary history and wide geographic range. We might also predict an unusual amount of variation in our T. rex samples because, if this one species was undergoing such a transformation across three decades of growth, it would have transitioned through a large number of “morphs”. Coupled with distortion caused by fossilisation and the dusting of individual variation we’d expect among a reasonable sample of biological entities and we're going to find a lot of variation among our Tyrannosaurus fossils. It’s an interesting idea that explains a lot of weirdness around our T. rex sample, perhaps more parsimoniously than greater taxonomic granularity or sexual dimorphism. But on those topics…
Graphic summary of select skulls referred to T. rex and T. "x"by Larson (2008). As above, this is not a comprehensive assessment, but shows where important, name-bearing specimens end up in Larson's scheme.
Developing in parallel to this “everything is Tyrannosaurus” interpretation were research projects that argued to split T. rex apart. The drip of new Tyrannosaurus specimens that started in the 1960s and ‘70s had turned into a relative torrent in the 1990s, such that researchers of the 2000s had dozens of T. rex specimens to examine. This was a large enough sample that we had some hope of teasing more obvious trends or patterns out from the statistical noise of individual variation and we might, finally, get some perspective on whether T. rex was one species or several.
In 2008, Pete Larson specifically addressed this issue (Larson 2008a). He noted that some adult Tyrannosaurus skeletons can be much stockier and more heavily built than others and ascribed this to sexual dimorphism for a variety of reasons, most of which are now doubted. But he also highlighted variation in cranial pneumatic features, tooth counts and the number of small "incisiform" teeth at the front of their lower jaw. These, Larson argued, were more likely to be taxonomic differences. Only a few specimens possessed a full suite of distinctive features that separated them from T. rex proper, including the giant skull MOR 008, the privately-owned “Sampson” skeleton, and the iconic AMNH 5027. Larson set these aside from the T. rex hypodigm but erred on the side of caution by not establishing a new taxon: instead, he gave this potential second Tyrannosaurus species a nickname: Tyrannosaurus “x”.
The holotype dentary of T. rex shows one small "incisiform" tooth in this photo presented by Larson (2008). The presence or absence of a second small tooth at the front of the lower jaw has been suggested to denote a distinct Tyrannosaurus species, but different authors disagree over the true variation of this feature.
The decade following Larson’s proposal has not provided much support for his suggested split in Tyrannosaurus specimens. In part, this reflects weak statistical support for the alleged taxonomic and sexual morphotypes. Joshua Smith’s (2005) assessment of T. rex dental characteristics and their utility in systematics can be seen as a blind test of size variation in the second dentary tooth, for instance, but failed to find significant characteristics of this in examined specimens. Mallon (2017) specifically looked for evidence of Tyrannosaurus sexual dimorphism along the parameters outlined by Larson but failed to find the sort of bimodal character distribution that would, in theory, result from differentiated sexes. More recently, Thomas Carr’s gigantic 2020 ontogenetic analysis of 44 T. rex specimens also found no taxonomic significance to Larson’s proposed dental and skull characters, nor did it recover evidence for sexual dimorphism. Moreover, it also failed to cluster the specimens ringfenced by Larson as T. "x". The size of Carr's analysis really has to be emphasised; by cataloguing up to 1850 features in 44 specimens, this investigation is, by far, the most in-depth analysis of anatomical variation within Tyrannosaurus (and maybe any dinosaur), and its failure to recover conclusions similar to those of Larson's 2008 study does not look good for the T. "x"hypothesis. Indeed, the scope and depth of Carr's investigation sets a high bar to clear for any different interpretation of T. rex variation, and that brings us to…
…Paul et al. (2022): the “mini-revolution for the dinosaur long universally called T. rex”
The suggested T. rex taxonomy of Paul and colleagues (2022) and its potential fallout from collisions with older schemes. Again, this is not a comprehensive breakdown of which T. rex specimens go where in this taxonomy, it only shows the treatment of key, named specimens.
As will now be clear, a finger has been hovering over the big red button marked “new Tyrannosaurus species” for decades now, and someone, somewhen, was going to push it eventually. In this context we can see Paul et al. (2022) as simply being the team that took the plunge, offering to give yet another interpretation of Tyrannosaurus ;taxonomy by splitting T. imperator and T. regina from T. rex. And as will also now be clear to anyone familiar with Paul and colleagues’ arguments, there is a certain historic familiarity to some of their ideas: the characters used are very similar to those used by Larson for characterising T. "x"; specifically, regina, rex and imperator are distinguished by features of skeletal robustness and the number of smaller incisiform teeth in the lower jaw. To be fair, how these are quantified and measured is slightly different in Paul et al., but there's undeniable conceptual overlap. I suspect this is one reason the paper has not received the most enthusiastic response among tyrant dinosaur experts, who will not only have experienced a sense of deja vu from the concept of new Tyrannosaurus species but also immediately thought of those analyses which have already investigated and dismissed the taxonomic potential of robustness and dentary incisiforms. And, OK, we need to be careful not to be arrogant or dismissive in these considerations: it’s really easy to say “other analyses didn’t find this result, so you’re probably wrong”, but if a new paper is essentially arguing points that have already been shown as doubtful, it’s fair to point to the same studies that dismantled those ideas the first time around.
And it’s not only in character choice that the past might catch up with the conclusions drawn by Paul et al. As outlined above, T. rex taxonomy has enough history and complications under its skin that it’s hard to create, as Paul et al. attempt, a totally fresh Tyrannosaurus classification without running into leftovers from older proposals. For instance, Paul et al. refer the holotype of Dinotyrannus megagracilis to their new species T. regina, arguing that the megagracilis type is an incomplete juvenile specimen and thus a nomen dubium, and therefore unsuited to be the holotype for their new species. But, unless my understanding of ICZN rules is totally wrong, I’m not sure that’s how nomenclatural rules will shake out. For megagracilis to be identified as regina it must show at least some of the features Paul et al. assert are unique to T. regina; this being the case, it (theoretically) has features distinguishing it from T. rex as well. If a specimen can be identified to species level it cannot be a nomen dubium, and this should would allow megagracilis to re-enter the field as a valid, non-T. rex species. And if megagracilis is the same species as regina, then megagracilis has nomenclatural priority by 34 years: the "Tyrant Queen" becomes… "Tyrant Big Skinny". Note that megagracilis is also a Paul (1988) taxon, so it’s regina’s own senior author complicating the attempted, and admittedly pretty neat, emperor-king-queen naming scheme.
The imperator name is potentially at risk from an older name too, as highlighted by tyrannosaur expert/palaeontology social media machine Thomas Holtz in the New York Times. The holotype of Nanotyrannus lancensis stems from the lower portion of the Hell Creek Formation where imperator, according to Paul et al., is said to have existed independently from rex and regina. Given that the lancensis type specimen is clearly a juvenile animal, and most probably Tyrannosaurus (Carr 1999; Woodward et al. 2020), parsimony suggests it should be the same species as T. imperator. Paul et al. (2022) are non-committal about Nanotyrannus as a separate genus, noting its questionable status but not coming down on either side of the debate around its validity. But whether we regard the generic name as warranted or not, if Nanotyrannus is the juvenile of a stratigraphically older species of Tyrannosaurus, lancensis would have nomenclatural priority over imperator. The Tyrant Emperor would become humdrum T. lancensis.
Variation within Tyrannosaurus as illustrated by Paul et al. (2022): how many species do you see? What's worth noting in this graphic is that two specimens (B, RSM 2523.8; and F, AMNH 5027) are not referred with confidence to any Tyrannosaurus species by Paul and colleagues, which seems remarkable given their relative completeness.
It’s also interesting to see how Paul and colleagues’ conclusions compare to those of previous workers. Of note are that specimens highlighted by others as potential new Tyrannosaurus taxa — such as AMNH 5027, the Texan maxilla — are considered indeterminable by Paul et al., and that Larson (2008a), despite utilising the same dentary tooth character, retained the holotypes of regina and imperator in T. rex. There’s nothing wrong with different projects drawing contrasting taxonomic conclusions, of course, but this shows that attributes as basic as specimen tooth size are being interpreted very differently by different authors. This demonstrates the difficulty, and perhaps the degree of subjectivity, associated with carving our Tyrannosaurus dataset into multiple species. Perhaps the clusters of specimens we're identifying as distinct "species" are strongly influenced by our own opinions on what features are taxonomically important, and also how we choose to measure and characterise those features? The discussion of Tyrannosaurus dentary incisiform size by Paul et al. is telling in this regard, highlighting the existence of “borderline” specimens that could be interpreted one way or another. Differing takes on such specimens probably account for the varied views proposed by different researchers.
The end? Probably not...
The take-home here is a simple one: T. rex is a tricky animal to split into multiple taxa, but the problem is not because researchers are wedded to the idea of there being one species of this animal. To the contrary, exactly the opposite is true: people have been trying to blow T. rex into smaller taxonomic units for half a century! Rather, it's because what seems like a straightforward job of naming a few new tyrannosaur species is complicated by the enormously variable nature of the T. rex dataset and also the lesser-known, but still important, taxonomic considerations of past workers. I admit to seeing Paul and colleagues’ work as less of a “mini-revolution” than the latest contribution in a decades-long conversation about the variation apparent in Tyrannosaurus specimens. Their work is distinguished, of course, by finally erecting new Tyrannosaurus species after 50 years of researchers tossing this idea around, and in this respect, Paul et al. have moved the conversation on. But in doing this, a greater spotlight is now shone on past attempts to investigate Tyrannosaurus variation, including the legitimacy of species previously sunk into T. rex as well as the value of anatomical characters already of doubtful value in dividing up the T. rex sample. It’s the response to these that will determine how far our understanding of Tyrannosaurus diversity has been moved by these new proposals.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored through Patreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusive Patreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Bakker, R. T., & Williams, M. (1988). Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. Hunteria, 1(5), 1.
Brochu, C. A. (2003). Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Journal of Vertebrate Paleontology, 22(sup4), 1-138.
Brusatte, S. L., & Carr, T. D. (2016). The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Scientific Reports, 6(1), 1-8.
Carpenter, K. (1990). Variation in Tyrannosaurus rex. Dinosaur systematics: perspectives and approaches, 141-145.
Carr, T. D. (1999). Craniofacial ontogeny in tyrannosauridae (Dinosauria, Coelurosauria). Journal of vertebrate Paleontology, 19(3), 497-520.
Carr, T. D. (2020). A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence. PeerJ, 8, e9192.
Carr, T. D., & Williamson, T. E. (2004). Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America. Zoological Journal of the Linnean Society, 142(4), 479-523.
Currie, P. J. (2003). Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the upper Cretaceous of North America and Asia. Canadian Journal of Earth Sciences, 40(4), 651-665.
Gilmore, C. W. (1946). New carnivorous dinosaur from the Lance Formation of Montana. Smithsonian Miscellaneous Collections 106 (13), 1-19.
Glut, D. F. (1997). Dinosaurs: the Encyclopedia. McFarland and Company Inc.
Holtz Jr, T. R. (2021). Theropod guild structure and the tyrannosaurid niche assimilation hypothesis: implications for predatory dinosaur macroecology and ontogeny in later Late Cretaceous Asiamerica. Canadian Journal of Earth Sciences, 58(9), 778-795.
Larson, P. (2008a). Variation and sexual dimorphism in Tyrannosaurus rex. In: Larson, P. & Carpenter, K. (eds.) Tyrannosaurus rex: the Tyrant King. Indiana University Press, 103-130.
Larson, N. L., (2008b). One hundred years of Tyrannosaurus rex: the skeletons. In: Larson, P. & Carpenter, K. (eds.) Tyrannosaurus rex: the Tyrant King. Indiana University Press, 1-56.
Larson, P. (2013). The case for Nanotyrannus. In: Parrish, J. M., Molnar, R. E., Currie, P. J. and Koppelhus, E. B. Tyrannosaurid paleobiology. Indiana University Press, 15-53.
Lawson, D. A. (1976). Tyrannosaurus and Torosaurus, Masstrichtian dinosaurs from Trans-Pecos, Texas. Journal of Paleontology, 50 (1), 158-164.
Mallon, J. C. (2017). Recognizing sexual dimorphism in the fossil record: lessons from nonavian dinosaurs. Paleobiology, 43(3), 495-507.
Molnar, R. E. (1991). The cranial morphology of Tyrannosaurus rex. Palaeontographica Abteilung A, 137-176.
Olshevsky, G., & Ford, T. L. (1995). The origin and evolution of the Tyrannosauridae, part 1. Dino Frontline, 9: 92–119.
Osborn, H. (1905). Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of American Museum of Natural History, 21, 259-265.
Osborn, H. F. (1906). Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication). Bulletin of the American Museum of Natural History, 22, 281-296.
Osborn, H. F. (1917). Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bulletin of the American Museum of Natural History, 35, 733-771.
Paul, G. S. (1988). Predatory dinosaurs of the world: a complete illustrated guide. Simon & Schuster.
Paul, G. S., Persons, W. S., & Van Raalte, J. (2022). The Tyrant Lizard King, Queen and Emperor: multiple lines of morphological and stratigraphic evidence support subtle evolution and probable speciation within the North American genus Tyrannosaurus. Evolutionary Biology, 1-24.
Rozhdestvensky, A. K. (1965). Growth changes in Asian dinosaurs and some problems of their taxonomy. Paleontologicheskii Zhurnal, 1965, 95-100.
Russell, D. A. (1970). Tyrannosaurs from the Late Cretaceous of western Canada. National Museum of Natural Sciences, Publications, in Paleontology, 1, 1-34.
Sampson, S. D., & Loewen, M. A. (2005). Tyrannosaurus rex from the Upper Cretaceous (Maastrichtian) North Horn Formation of Utah: biogeographic and paleoecologic implications. Journal of Vertebrate Paleontology, 25(2), 469-472.
Smith, J. B. (2005). Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions. Journal of Vertebrate Paleontology, 25(4), 865-887.
Woodward, H. N., Tremaine, K., Williams, S. A., Zanno, L. E., Horner, J. R., & Myhrvold, N. (2020). Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus. Science Advances, 6(1), eaax6250.
Behold, the cover of The Art and Science of the Crystal Palace Dinosaurs, available for preorder now, and on general sale next month!
Next month sees the publication of what might be one of the more important projects I’ve ever been involved with: a new book, The Art and Science of the Crystal Palace Dinosaurs, co-authored with Ellinor Michel and published by Crowood Press. This is a large, richly illustrated hardback that, as the title suggests, discusses the creation, scientific content, artistry and historic legacy of the world-famous Crystal Palace Dinosaurs, a story we tell in lots of detail and with hundreds of photos, illustrations and diagrams, both vintage and modern.
The good news is that, as of a few days ago, the book became officially available for preorders so palaeoart and history of science aficionados can start bagging copies for delivery next month. The exact publication date has been hard to pin down because of the many global incidents disrupting shipping and transportation, but mid-May is looking like the point when it will be available. You’ll be able to pick it up from just about wherever books are sold, but, with apologies to North American readers, you’re going to have to wait a bit longer for your release as you’re on a different distribution network to us here in the UK (that’s not to say you can’t order it from Europe and have it shipped over to you, of course). The cover price is £30 and, as we’ll detail more below, every sale directly contributes towards the care and maintenance of the Dinosaurs themselves.
With preorders now being taken, we can finally start to talk about our book more openly. The Crystal Palace Dinosaurs can be described, without exaggeration, as some of the most famous pieces of palaeoart in the world and they are true mainstays of dinosaur books and documentaries. They encompass a series of Victorian prehistoric animal sculptures and recreated geological features based in Penge, in the southeastern suburbs of London. The most famous components of the site are the prehistoric animal models, which were built between 1852 and 1855 by their chief architect and artist, Benjamin Waterhouse Hawkins, ostensibly under the watchful eye of Victorian palaeontological mastermind Richard Owen. Much of the original site is still with us today and can be visited freely if you want a glimpse of what cutting-edge palaeontology looked like in the early 1850s. Of the 30 sculptures still standing, the four dinosaurs, representing Iguanodon, Megalosaurus and Hylaeosaurus, are genuine icons of 19th century palaeoart, and a large amount of effort has been exerted over the years to keep the site in fair condition (though read on).
The face that launched this particular ship: the broken Crystal Palace Megalosaurus, as photographed by the Friends of Crystal Palace Dinosaurs in May 2020. As this image shows, the conservation risk to the site is very real and a sense of wanting to do something to help is what got this book rolling.
The origins of our book give a pretty good idea of our aims and hopes for this project. In May 2020 the jaws of the Crystal Palace Megalosaurus were severely damaged in a mysterious incident: the cause is assumed to be final succumbence to weathering or simply vandalism, or a combination thereof. Both, sadly, are common agents of deterioration at the site. The photos of the damaged sculpture shared online were pretty disheartening and, having been working with the Friends of Crystal Palace Dinosaurs to augment their website (you may remember a series of blogposts I wrote related to this project from 2019: part 1, 2, 3 and 4), I wondered if turning my notes and illustrations into a book that we could sell to benefit the sculptures was a good idea. I floated this to Ellinor, who's chair of the Friends of Crystal Palace Dinosaurs, and we then approached Crowood, who you may know from my Palaeoartist’s Handbookand Emily Willoughby’s Drawing and Painting Dinosaurs, to make the project a reality. The pitch was to analyse the Crystal Palace Dinosaurs as an enormous palaeoart project where we looked at their conceptualisation, construction and legacy in context with 19th century palaeontology and palaeoartistry while also, in addition, creating a book that would directly benefit the Dinosaurs themselves. To that end, neither Ellinor nor I have received a penny for producing it: all the usual advances and royalties that go to book authors are instead being donated to the Friends of Crystal Palace Dinosaurs to further their efforts in understanding and maintaining the Dinosaurs and their home.
We finished writing the book in August 2021 and, I must admit, it was a lot more work than I initially anticipated. This not only reflected the complications of writing a book during Covid-induced national lockdowns but also the volume of material to discuss. Ellinor and I are far from the first scholars to write about the Crystal Palace Dinosaurs, with notable contributions to the literature having been made by Martin Rudwick (1992), Peter Doyle (Doyle and Robinson 1993, 1995; Doyle 2008), Jim Secord (2004), Gowan Dawson (2016), Valerie Bramell and Bob Peck (2008), and Steve McCarthy and Mick Gilbert (1994). Our palaeoart-focus, aided by the modern searchability of digital archives, meant we were able to unearth a lot of obscure details about the Dinosaurs not mentioned elsewhere. My initial thoughts that this would be a quick and straightforward project — perhaps not much more than expanding and stapling my blog series together and writing a few discussion chapters — quickly evaporated when the amount of information to sift through and analyse became apparent. The book ballooned by 20,000 words from my initial projection and it was still a squeeze to get everything in. My somewhat immodest view is that we’ve compiled a new, richer synthesis about the Crystal Palace Dinosaurs through both our own findings as well as coalescing important points raised by other recent authors into our text; in doing so, we’re helping to establish a deeper narrative about this familiar, but still only partially understood site.
A sneak peek at one of the early chapters of the book, focusing on the 1853 New Year's Eve banquet held inside the clay mould of the standing Iguanodon.
Wrangling the story of the Crystal Palace into some sort of order created a book of three parts and 13 chapters:
Part 1. Islands covered by strange figures
1. Historic prehistory in South London
2. Ancient worlds through a Victorian lens: planning the Geological Court
3. Bricks, iron and tiles: rebuilding the past
Part 2. Animals long since extinct
4. The sculptures: Mammals
5. The sculptures: Mosasaurus hoffmanni
6. The sculptures: Flying reptiles
7. The sculptures: Dinosaurs
8. The sculptures: “Teleosaurus chapmani”
9. The sculptures: Enaliosaurs
10. The sculptures: “Labyrinthodon”
11. The sculptures: Dicynodon
Part 3. A difficult and, perhaps, too bold, attempt
12. The reception and legacy of the Geological Court
13. Past becomes future: the conservation of the Geological Court
You can gauge a lot about the book from that chapter list but, to get a superior sense of what we cover, let’s go into a little more detail.
Part 1. Islands covered by strange figures
The first section serves as an introduction to the world of the Crystal Palace Dinosaurs by discussing the principles and individuals behind their construction as well as the building of the site itself. Very quickly within the narrative, we establish that the label “Crystal Palace Dinosaurs” is not always useful or apt because it omits the major structures that accompanied Hawkins’ palaeontological sculptures: the Geological Illustrations. These are a series of reconstructed geological outcrops that extend around the Dinosaurs' landscape, being created either by importing tonnes of rocks chosen for their age and fossil content from sites around the UK or, alternatively, by recreating complex sedimentary strata using building materials. These rocks were not distributed haphazardly around the park, either, but in realistic stratigraphic congruence: in other words, they are in proper geochronological order, such that Triassic rocks are next to Jurassic rocks, which are then next to Cretaceous rocks, which are then next to "Tertiary" rocks and so on, and the palaeontological sculptures were placed around these in an appropriate geological context. The almost uninterrupted sequence (there are deliberately no Permian rocks) allows visitors to walk continuously from the Devonian to the Quaternary, seeing signature rock types and fossil species along the way.
Our map summarising the full extent, both planned and actual, of the Geological Court. Note the complexity of the geology as well as large numbers of missing models, denoted by red graphics and text, and the extent of never-realised models on the Tertiary Island.
Mapping the strata as real geological features reveals a very sophisticated and complex arrangement of manufactured geology, even incorporating simulated unconformities (missing rock layers, which account for the absence of Permian features) and faults to condense much of the geological column, as it was known in the 1850s, into a small area (Doyle and Robinson 1993). The economic importance of geology to Victorian culture was highlighted with not only a partly artificial coal seam (real coal, partially fake rocks) but a motherflippin’ artificial mine and cave, some 20 m long, within which manufactured stalactites and floatstones, along with mining tools, gave visitors the experience of traversing a real lead mine. The unsung heroes of this forgotten aspect of the park were geologists David Thomas Ansted and James Campell, and we suggest that they need as much recognition as Hawkins or Owen for their contributions to this project. Reflecting this, we mostly refer to the “Geological Court” instead of “Crystal Palace Dinosaurs” throughout the text. This was the name originally given to Crystal Palace’s combined geological and palaeontological display and better encapsulates the full extent of the project. The Geological Illustrations receive a lot of attention in our book, for which we need to tip our hat to Peter Doyle for laying critical interpretive groundwork that we could build on (Doyle and Robinson 1993, 1995; Doyle 2008).
Today located far away from public footpaths is the Carboniferous Mountain Limestone, which contains an artificial cave and lead mine. This is one of the most complex of all the Geological Illustrations and it's also huge: the original Mountain Limestone "outcrop" represented 90 tonnes of real Carboniferous limestone imported from Matlock, in the UK Midlands. This was removed in the mid-20th century so the Mountain Limestone you see now (including in this photo) is a reconstruction from the early 2000s. The cave is original, but is now half-filled with sediment and inaccessible to the public.
Our opening chapters also offer a deep dive into the construction of the models themselves, both in terms of the palaeoartistic principles employed by Hawkins as well as the physical building process. We speak a lot about the importance of 19th century “anatomical correlation” in predicting the life appearance of fossil animals from scrappy bones (Dawson 2016) and analysed photographs and illustrations of the models under construction — some familiar, others less so — to obtain new details of how these often gigantic creations* were assembled. It seems there was no one method behind their realisation, but a mix of techniques that probably depended on practical considerations as well as the availability and cost of materials. The Megatherium was unqiuely carved from blocks of limestone (Doyle 2008) while most of the models were composites of bricks, concrete and metal that could be moved about the site on carts and trollies. The dinosaurs were constructed more like houses, adding bricks, mortar and concrete around deep foundations and enormous iron frameworks (Hawkins 1854). The construction of the Court took place under the eager eyes of international media, and we also use these early chapters to review this interest in the Geological Court. Part of this discussion focuses on the famous 1853 New Year’s Eve banquet in one of the clay Iguanodon moulds. This event has been retold so often that a certain amount of truth has leaked from the original story but, using newspaper accounts and archive material, we think we’ve managed to tidy up what truly happened and answer obvious questions, such as how several dozen people squeezed into a very large, but not enormous dinosaur belly (spoilers: there was an adjoining table, so not everyone sat within the Iguanodon itself. See this Twitter thread if you’d like to know more).
*And I do mean “gigantic”: the Megalosaurus is 12 m long and the Iguanodon and Temnodontosaurus are not much smaller. Photos don’t convey how seriously big some of these sculptures are, and another novelty of our book is providing basic measurements of each one, an obvious aspect to report but, to our knowledge, unrecorded until now. My boots are still drying out from wading through the waters surrounding the marine reptiles.
Part 2. Animals long since extinct
The middle of the book is a sculpture-by-sculpture analysis of the palaeontological creations, comprising the mammals, the various marine reptiles, the pterosaurs, dinosaurs, amphibians and Dicynodon. This is the longest section of the book and perhaps the main draw for palaeoart fans. Alas, we still don’t have much insight into Hawkins’ original plans for his sculptures — no relevant notebooks, sketches or correspondence to this effect are known anywhere — but efforts were made to “reverse engineer” the palaeoartistry of each model by comparing what Hawkins produced against palaeontological science of the 1850s. This was similar to the approach I used in my Crystal Palace Dinosaurs blog posts but we go way beyond the details discussed in those articles. Each sculpture is given a photographic montage to show features of interest as well as a diagram showing what fossil specimens or modern animals likely referenced each body part.
An example of the breakdowns given to each restored species at Crystal Palace: what fossils were available to inform Hawkins' restorations? Here, we see how the Iguanodon is really a hodgepodge of different iguanodont material, and not a reconstruction of a single species.
Analysing the sculptures at this level allowed us to break down their “real” identities, and we can see today that many were chimeric mixes of different species. "Labyrinthodon pachygnathus", for instance, can be considered an early attempt at reconstructing the (possible) ctenosauriscid (those neat sail-backed croc-line archosaurs) Bromsgrovia rather than a prehistoric amphibian, and the known reference specimens for Iguanodon do not include any “true” Iguanodon material in the sense that we recognise it now. It’s actually difficult to know what to classify the Iguanodon as in a modern sense, as at least two, and maybe three iguanodont species were factored into its restored form. If we force the issue, Barilium dawsoni, a large iguanodont that was used to establish the size of the Iguanodon sculptures and other components of its anatomy, is perhaps the most dominant species used in the build, so maybe that’s the closest we have to the “true” identity of these sculptures.
Assessing the fossil composition of Hawkins' recreations allowed us to ally species to the Crystal Palace project that are not normally associated with it. They include the possible ctenosauriscid Bromsgrovia walkeri, which is too poorly known to restore itself, but might have resembled Arizonasaurus babbitti. This is a nice reminder that the British fossil record contains a lot of remarkable animals, even if some of their fossils are less than exemplary.
We also compare each species with modern interpretations of the same taxa, and I used these sections as an excuse to sneak in some of my own artwork to show 21st views of the Crystal Palace species. These are all-new restorations rather than recycled images from my 2019 blog posts, and some feature fun callbacks to classic Hawkins imagery. It’s easy to be a little blasé about some of the Crystal Palace taxa because many are well-trodden palaeoart subjects, like Megalosaurus, Megaloceros, Iguanodon etc., but the reality of some of the animals restored for the Geological Court is pretty wild. Leptonectes, for instance, with its grumpy face and massive pectoral fins, is definitely an unusual ichthyosaur, while Cimoliopterus — the animal behind the Chalk pterosaurs — belongs to the long-winged, giant-headed pterosaur group Ornithocheiromorpha. Among everything else, this book was a nice opportunity to draw attention to some lesser-known British fossils that, owing to being poorly preserved, are often overlooked.
I created something like 30 new paintings for The Art and Science of the Crystal Palace Dinosaurs, including this image of the uppermost Triassic ichthyosaur Leptonectes tenuirostris. This reality of this frowny-faced, giant-flippered ichthyosaur is quite different to what was restored at Crystal Palace, where it looks a lot more conventional.
Looking at the sculptures in such detail allowed us to write at length about how excellently executed they are, from their depicted musculature and fine anatomical detailing to their considered behavioural depictions. Hawkins was really ahead of his time by creating palaeoart that showed plausible, realistic-looking animals rather than, as was then common, either super-conservative reconstructions with minimal detailing or, more commonly, fantastic restorations that have more in common with mythology than zoology. Interestingly, we did find evidence that Hawkins considered more aggressive and dynamic posing for at least some of his models before settling on their more sedate poses.
We also compared Hawkins' work to Owen’s publications to gauge how closely Hawkins was working with his consultant. The large number of deviations we found is, we argue, further evidence of Owen being a pretty useless consultant. Owen's contributions to the project have been enormously overstated, with records showing that he neglected to visit the construction site or Hawkins' workshed outside of a handful of instances, and that was also largely ignorant of the appearance of the sculptures until he saw them completed and installed in the park grounds (Secord 2004; Dawson 2016). That Hawkins made some errors Owen could have corrected is entirely consistent with this narrative. Owen also loses points in our dissection of his 1854 Geological Court guidebook, which is often inconsistent with the content of the site itself. Indeed, Owen’s slim overview of the site didn't mention major components of the display, including the bulk of the Geological Illustrations and mammal sculptures. Owen doesn’t quite warrant writing out of the history of the Geological Court, but he definitely needs recasting as a peripheral character who did little to help Hawkins and the Crystal Palace Company accomplish their goals.
The missing paleontological sculptures of Crystal Palace: two pterosaurs, three "Anoplotherium gracile", one Palaeotherium magnum and one female Megaloceros (red arrow). Note the different face on "Palaeotherium minus" and the real antlers on Megaloceros; both have since been replaced with substitutes that no longer resemble the originals. The photographs in this image were kindly provided by the Crystal Palace Foundation.
The final, and perhaps most significant, component of these chapters I want to mention is their discussions of missing models. Exactly how many models and the number of different species they represent has been variably interpreted (e.g. Doyle and Robinson 1993; McCarthy and Gilbert 1994) because of the complex and tumultuous history of certain sculptures. Some have been moved about, mislabelled or even been removed from the site over the last 170 years, leaving researchers to draw different conclusions about what’s left in the park today, and what was there originally. We attempted to get definitive numbers on both counts and concluded that our modern Geological Court holds 30 sculptures of 21 species. In 1855 however, when the Court was at its most complete, it had 37 sculptures and 24 species. That’s a higher count than any previous calculation, but one we’re confident about thanks to data from historic guidebooks, illustrations and photographs. The seven lost models include a fourth Megaloceros (specifically, another model of a reclined doe), the two Jurassic pterosaurs, the large Palaeotherium species P. magnum and, finally, three models of “Anoplotherium gracile”.
The "Megaloceros fawn", which we reidentify as a different animal entirely: "Anoplotherium gracile" or, in modern terminology, Xiphodon gracilis. As explored more below, there is no evidence that this animal represents a juvenile Megaloceros but plenty of evidence to tie it to Xiphodon. How and why this sculpture became associated with the Giant Deer display is not currently understood.
The existence of the latter three sculptures is worth going into a little more because it’s one of our biggest discoveries and ties into another important conclusion. If you’ve visited the Crystal Palace Dinosaurs you’ll know that a small Megaloceros fawn sits close to the three surviving Giant Deer adults in the Quaternary end of the Court. We present evidence that this, in fact, is not a juvenile deer at all, but the sole survivor of a group of four “Anoplotherium gracile”. Known as Xiphodon gracilis today, the presence of this guanaco-like Eocene species among the Crystal Palace fauna has long confused researchers because, despite being mentioned in several guidebooks, it was thought that no obviously Xiphodon-like animals were known at the site, nor were any witnessed in familiar vintage photos or illustrations. Some authors have attempted to reconcile these facts with a more gracile Anoplotherium commune sculpture, assuming that this represented Xiphodon (Doyle and Robinson 1993; McCarthy and Gilbert 1994), but I’ve never found this interpretation convincing. Even in the 1850s scholars knew that A. commune and Xiphodon contrasted in size, build and proportions. The “fawn”, however, is a dead-ringer for historic takes on Xiphodon anatomy, and an image of Hawkins’ workshed shows this same sculpture with three others of the same species (above). Conversely, we found no evidence whatsoever of a Megaloceros fawn existing at the site before modern times. Putting these pieces together, it looks like the blank spots in the history of the displays are large enough to both obscure the loss of three Xiphodon sculptures and also hide the relocation of the surviving Xiphodon alongside the Giant Deer, where it has masqueraded as a juvenile Megaloceros for the entirety of living memory. I admit to finding this as worrying as I do exciting. Discovering not just one Xiphodon but records of four is very cool, but it also shows how enormous the holes in our knowledge of the Geological Court are. If something as fundamental as the existence of a whole set of sculptures can go virtually unrecorded, what else are we missing?
Part 3. A difficult and, perhaps, too bold, attempt
The last section contains two chapters, one on the complex legacy of the Geological Court and another on its constant battles with degradation. The chapter on the post-development history of the Crystal Palace Dinosaurs is one of the largest in the book and attempts to make sense of a complicated story. It’s fair to say that they were neither the major successes nor major failures they have been portrayed as by different authors, and reactions to the Geological Court have fluctuated enormously in the last 170 years. Early scientific views, for example, were very hostile. Hawkins’ rapidly-dating sculptures vindicated palaeoart sceptics who saw full, elaborate restorations of extinct life as premature and, as has been noted by others (Secord 2004; Nieuwland 2019), the Geological Court seems to have lessened appetites for palaeoart among many, perhaps most, 19th century palaeontologists. It took decades for more sympathetic views to develop among academics (e.g. Hutchinson 1893; Becker 1911) and for palaeoart to regain its early 19th century mojo. Among the public, the Geological Court was a source of equal parts fascination and confusion, as the displays — which lacked any sort of signage or explanation, as per Crystal Palace Company policy — represented content too far from general knowledge for lay audiences to grasp their significance. Some gathered that they were looking at animals that existed before humans (Martineau 1854), but others thought they were grossly-distorted sculptures of living species, or even attempts to show the dangers of intoxication (Owen 1894). As well as cataloguing this diversity of opinion, we also cover the cancellation of the Geological Court’s development, the unrealised models and geological components (in addition to at least a dozen more mammals and birds, planned works included additional Cambrian, Silurian and “Tertiary” Geological Illustrations), vintage Crystal Palace Dinosaur merchandise, and the career impacts of the project for Hawkins and Owen.
A selection of dinosaur palaeoartworks produced after Crystal Palace. It took several decades for scientists to start regularly commissioning artwork of new fossil animals, allowing Hawkinsian dinosaurs to linger in paleoartworks until the late 19th century, as in the lower right images. Novel reconstructions of upright dinosaurs were produced as early as the 1860s (top images), but were rare until palaeoart got its mojo back in the 1890s. Was the scientific backlash against the Crystal Palace Dinosaurs responsible for this dearth of new artworks? There is certainly circumstantial evidence supporting this view.
All this leads to our final chapter, the inevitably stern-faced discussion of the site’s ongoing conservation and maintenance issues. It would have been nice to end the book on a more positive note but it would have been misleading to portray anything other than the truth: that our collective efforts to keep the Geological Court in good condition have not been exemplar and, unless this changes, the long-term outlook for the site isn’t great. We review the patchy conservation history of the site and highlight that no consistent approach to maintenance has ever been followed: repair work has really been a series of occasional interventions, not a routine, regular event. Even more amazingly, outright destruction of some components were justified in the mid-20th century to make way for other Park developments. We can’t be sure, but we think these events were likely responsible for the removal or destruction of the missing palaeontological sculptures. These issues have persisted to modern times and it's a sad fact that the current appearance of the site we're attempting to celebrate is rather sorry. We often had to source photos from the last decade to illustrate the displays in a more intact, less overgrown condition.
A shot from my last visit to the Geological Court in July 2021, showing the extent of overgrowth and degradation among some of the marine reptiles. To give a sense of the scale of the vegetation in this photo, that middle ichthyosaur is one of the largest models at the site: the c. 12 m long Temnodontosaurus. The broken jaw of Ichthyosaurus communis, which you can see in the background, was a recent incident which has now been fixed. Needless to say, allowing the displays to get to this state is pretty criminal, and without a significant, long-term change to maintenance and conservation approaches, we risk losing the site forever.
Thankfully, the Crystal Palace Dinosaurs are now protected against further wanton development by having attained Grade 1 listed status. This recognises the Geological Court as having exceptional historic interest and brings a level of care and protection from Historic England, the public body that manages places of national importance to English heritage. Much is hoped from this relatively recent development and projects to save the site are underway, but we shouldn’t pretend there isn’t a lot of work to do. Behind the visibly crumbling displays are problems as fundamental as subsidence and complex internal damage caused by degradation of their metal frameworks. In addition to dramatic interventions to solve these problems, a regular management strategy to keep on top of perpetual conservation risks is also critical. Without interventions to halt plant growth on the displays and deter human trespassers, the Dinosaurs and Geological Illustrations will quickly fall back into disrepair. Ensuring that the Geological Court endures for another 170 years will not be easy or cheap, and requires a broad shift in how we value the site as a nation. Today, we’re perhaps at a critical point for deciding its future. If we don’t invest our energy and money now, we may be among the last generations to witness something approximating its original grandeur.
TL,DR: buy our book; learn cool things; save some dinosaurs.
And that, in a several thousand-word nutshell, is our book. But this post is really just a teaser of what we have to say: there is so much more to discuss around the Crystal Palace Dinosaurs that we struggled to get everything into this one tome. But for all this, there’s still plenty that we couldn’t write about, because there are enduring mysteries that we were unable to crack. What, for example, are the mysterious Wisbech Museum models of the Crystal Palace species? Do we really not have a single record for what happened to all those palaeontological sculptures, not even in some council development office somewhere? Why were so many models repaired with unsuitable replacement parts at some point in the 20th century? Will anyone, ever, find some of the original designs for the Geological Illustrations or extinct animal restorations? These, and other questions, are for future researchers to look into. For now, we’re happy to have moved the conversation along as far as we have, and to once again be shining the light on the conservation plight of a site unique in its significance to the history of science. We’re especially happy because all this will simultaneously help fund the Friends of Crystal Palace Dinosaurs and their work monitoring and maintaining the Geological Court. Check out The Art and Science of the Crystal Palace Dinosaurs if you want to appreciate the full awesomeness of this very special place.
And finally…
As a quick closing comment, I want to extend a quick personal thanks to a group of people who made this book possible: the folks who support my work at Patreon. Researching, writing and illustrating a book is a huge amount of work and the only way I could commit so much time to a charity project like this was through their monthly donations. So sincere thanks to everyone who supports me there: any positive impact this book has is, in part, related to your continued donations.
References
Becker, H. (1911). Alte und neue Rekonstruktionen ausgestorbener Tiere. Die Umschau.
Bramwell, V., & Peck, R. M. (2008). All in the bones: a biography of Benjamin Waterhouse Hawkins. Academy of Natural Sciences.
Dawson, G. (2016). Show me the bone: reconstructing prehistoric monsters in nineteenth-century Britain and America. University of Chicago Press.
Doyle, P. (2008). A vision of ‘deep time’: the ‘Geological Illustrations’ of Crystal Palace Park, London. In: Burek, C. V. & Prosser, C. D. (eds). The History of Geoconservation. Geological Society Special Publications, 300(1), 197-205.
Doyle, P., & Robinson, E. (1993). The Victorian ‘Geological Illustrations’ of Crystal Palace Park. Proceedings of the Geologists' Association, 104(3), 181-194.
Doyle, P., & Robinson, E. (1995). Report of a field meeting to Crystal Palace Park and West Norwood Cemetery, 11 December, 1993. Proceedings of the Geologists' Association, 106(1), 71-78.
Hawkins, B. W. (1854). On visual education as applied to geology, illustrated by diagrams and models of the geological restorations at the Crystal Palace. Journal of the Society of Arts (78): 443-449.
Hutchinson, N. H. (1893). Creatures of other days. Chapman & Hall
[Martineau, H.]. (1854). The Crystal Palace. Westminster Review, 62, 534-50.
McCarthy, S., & Gilbert, M. (1994). The Crystal Palace Dinosaurs: The story of the world's first prehistoric sculptures. Crystal Palace Foundation.
Nieuwland, I. (2019). American Dinosaur Abroad: A Cultural History of Carnegie's Plaster Diplodocus. University of Pittsburgh Press.
Owen, R. (1894). The Life of Richard Owen. J. Murray.
Rudwick, M. J. (1992). Scenes from deep time: early pictorial representations of the prehistoric world. University of Chicago Press.
Secord, J. A. (2004). Monsters at the crystal palace. In: de Chadarevian, S, & Hopwood, N. (eds). Models: the third dimension of science. Stanford University Press. 138-69 pp.
By now many readers will have seen the excellent, excellent BBC/Apple TV+ documentary Prehistoric Planet, a show being widely celebrated as the most significant televisual portrayal of dinosaurs since 1999’s seminal Walking with Dinosaurs. There’s a huge amount to say about the reaction to Prehistoric Planet as well as how it was put together. I’m proud to say I was involved as a concept artist and consultant but, for now, those of us involved with the show have been (understandably) asked not to give too much away. We can, nevertheless, use the programme as a jumping-off point to discuss some cool dinosaur science, and that’s what I want to briefly do today.
One of the most memorable scenes in Prehistoric Planet is the opener of the first episode, and you might be familiar with it even if you haven’t seen the full series. It’s the sequence featuring the swimming Tyrannosaurus and his juveniles crossing a small body of water to engage in some beach-related foraging. Part of this scene featured in a promotional teaser that dropped so many jaws earlier this year, where Tyrannosaurus juveniles hunted turtles on a beach. If you've somehow missed this, here it is.
It’s easy to buy these superbly rendered, fluffy, long-legged and big-eyed baby tyrants as 100% accurate takes on such animals because they look so real. But under their wireframes and digital rigging, these juvenile dinosaurs are based, in fact, on hypothetical reconstructions, not fossil skeletons. Despite the intense interest in Late Cretaceous North American fossil sites, hatchling tyrant dinosaurs are virtually unrepresented by fossils and the first genuine remains of such animals were only described in 2021, by Greg Funston et al. These important finds constitute a piece of lower jaw, a tooth, and a foot claw from embryonic individuals of Daspletosaurus and Albertosaurus. Each sheds a little light on the size and proportions of baby tyrannosaurs, but we’re still far, far away from a complete picture of their entire osteology. Thus, to date, the only way to restore the life appearance of baby tyrants with some degree of scientific rigour has been to extrapolate their proportions from older, larger individuals. We’ve been attempting this activity for at least 50 years and, even now, we struggle to make accurate, or at least statistically sound, models of baby tyrant appearance.
The first effort at predicting hatchling tyrant anatomy was published in Dale Russell’s 1970 paper on the tyrant dinosaurs of Canada. Russell predicted the proportions of a hatchling “Albertosaurus” libratus (=Gorgosaurus libratus today) by calculating scaling trajectories of larger tyrants and projecting them to an individual with a 100 mm long femur. The resultant image is one that might be familiar to many readers, as it has appeared in subsequent dinosaur books and papers. I certainly remember my own first encounter with it in David Norman’s influential 1988 Normanopedia Illustrated Encyclopaedia of Dinosaurs, as it looked very different to the pop-culture plush toy-like baby dinosaurs I was familiar with (hello, The Land Before Time et al.).
Dale Russell's hypothetical Gorgosaurus libratus hatchling, from Russell (1970).
Russel’s prediction shows baby tyrants as long-legged, long-tailed and small-bodied animals that were very different to their parents, a consequence of the pronounced allometry that shaped tyrannosaurs as they grew. The overall animal, he calculated, was dog sized: just over 75 cm from the snout to the end of the tail. Some aspects of Russell’s model, including the shapes of the bones themselves, were created by “juvenilising” the bone shapes of adult albertosaurines and, even today, it's a good-looking skeletal. It certainly matches what we see in other non-avian dinosaur embryos and hatchlings, which are generally leggy animals adapted for precocial, relatively independent early lives. It’s probably this skeleton that a lot of palaeoart and dinosaur documentaries have based their baby tyrants on as, all these years later, it's still one of the only published hatchling tyrannosaur skeletals available.
In 2003 another attempt to predict tyrannosaur hatchling proportions was made by Phil Currie. Armed with more specimens and thus more metric data, Currie was able to provide more informed tyrant scaling regimes and, in theory, give us a superior prediction of hatchling proportions than Russell’s effort of three decades prior. As before, his scaling was to an individual with a 100 mm long femur, and the result was, again, dog-sized. Currie didn't specify what tyrannosaur he was modelling specifically, so his data can be viewed as a generic tyrannosaurid, although albertosaurine scaling metrics were prioritised where they differed significantly from those of tyrannosaurines. Because Currie (2003) only presented the results in a data table and did not illustrate the resultant hypothetical hatchling, I took the liberty to restore the animal with his suggested proportions myself*. And, holy moly…
The stilt-legged tyrant hatchling, restored after the dimensions given in Currie (2003). As explored below, the data behind this reconstruction were identified as unbelievable in Currie's original paper: this is not what Phil Currie (or anyone else) seriously thinks juvenile tyrannosaurus looked like! But it is what models of tyrannosaur ontogeny, as understood in 2003, predicted for a tyrant of hatchling dimensions.
…those are some l-o-n-g legs. They’re slightly maddening to look at, in fact. Like it’s been teleported in from the Nightmare Before Christmas meets Prehistoric Planet Super Crossover Spectacular. While the rest of the animal isn’t too dissimilar to Russell’s baby gorgosaur, the tibia is 60% longer and the metatarsals 220% longer, creating a tiny dinosaur on stilts. The result is a theropod with leg proportions outside of anything known within Dinosauria: even long-legged birds, like stilts, secretary birds and seriemas, are being put to shame. Needless to say, if anyone seriously thought that this is what baby tyrant dinosaurs looked like we’d be considering all sorts of ecological and biomechanical implications. Were baby tyrants waders? Specialists of prairies and open habitats? Did they make money on the side at Mesozoic carnivals as stilt walkers? What the heck is going on?
*I had to take lengths of the vertebral column and manus from Russell (1970), as these were not modelled in Currie (2003).
But, of course, no-one does think that this is what hatchling tyrannosaurs looked like. The outlandish hindlimb proportions of this model were dismissed as unbelievable by Currie (2003) as quickly as they were introduced, and the whole exercise ended with a blunt cautionary note: “there are limitations to what can be done in extrapolating this data.” (Currie 2003, p .663). And before anyone asks, it's worth stressing that Currie's values were not errors or the outputs of wrongful methodology: they're simply what fell out of legitimate, perfectly normal scaling equations. The "limitations" Currie referred to are fundamental difficulties with scaling the proportions of extinct animals outside of observed ranges. Such issues include low sample sizes, our scaling curves being skewed by outliers and overall low confidence intervals, but it's unusual to talk about these as problems for scaling tiny animals. We're generally much more used to these frustrations when trying to scale smaller animals to the size of giant adults.
And strange as it seems, I admit to finding Currie’s stilt-limbed tyrant baby a more profound example of these issues than our usual subjects of scaling woes, like titanic sauropods or giant pterosaurs, because we know so much more about small dinosaur proportions. It's easy to spot these data as (probably) “wrong” because they're so out of the ordinary, but concerning proportions are harder to intuitively identify in animals beyond the size limits of anything ever witnessed by a person. It's a good reminder that any extrapolation is viewed as shaky from a mathematical perspective, and recalls Knut Schmidt-Nielsen's comments in his classic 1984 book Scaling: why is animal size so important: "unless we fully understand all the pertinent factors involved (and that is not likely to happen in a biological system), going beyond the limits of observation is not simply chancy, but outright perilous” (p. 25). It’s perhaps only when we see relatable, but obviously wrong-looking reconstructions like the one above that we’re confronted with this reality. And while we basically have no choice but to extrapolate and go beyond data limits when dealing with certain unknowns in the fossil record, it's good to come back to Earth from time to time and realise the unreliable nature of the mathematics we're using. So many extrapolations we're familiar and comfortable with might be just as crazy as that lanky-limbed hatchling, we just don't see them as obviously "wrong".
All this said, that stilt-legged tyrant baby still has some utility. While it probably doesn't genuinely reflect what baby tyrants looked like, it does serve as a great visualisation of the allometry expressed through tyrannosaur ontogeny. If we compare the stilt-legged model with a regular-old T. rex, for instance...
The mad stilt-legged hatchling vs. a large adult Tyrannosaurus, based on "Sue".
...we get a good sense of how tyrant bodies grew into their legs as they progressed from hatchling to adults. We can also see that the head length remained about proportionate to femur length and that the arms grew relatively slowly, appearing to "shrink" a bit en route to adulthood (Currie 2003). Of course, in detail, there's actually a mosaic of different allometries occurring through growth: the head doesn't lengthen proportionately, for instance, but it does deepen. The arms and legs don't really "shrink" as one unit, either, but actually at varying rates in different parts of each limb (Currie 2003). Because the stilt-legged hatchling is our understanding of tyrannosaur scaling regimes expressed to an absurd degree, the reality of tyrannosaur ontogeny probably wasn't quite as extreme as the graphic above suggests, but it might not be too far off. Both Russell's (1970) and Currie's (2003) data point to long-legged, shallow bodied tyrant hatchlings and that's the model we should generally follow, even for chunky, robust animals like T. rex and Tarbosaurus. As noted by Currie in the same 2003 paper, "allometric differences among mature specimens of different [tyrannosaurid] species are shown to be trivial when compared with the allometric differences associated with growth". The stilt-legged tyrant (and more recent work and studies on this same topic) makes that crystal clear.
Currie's paper is now almost 20 years old so, to wrap this up, one last question: have we made any progress in being able to reconstruct baby tyrants reliably in recent years? To be frank, no, we don't yet have the data needed to clear the problems outlined above. Things have moved ahead thanks to Greg Funston and colleagues (2021) who, finally, have been able to factor some embryonic tyrant fossil material into their scaling equations. These have generally pointed to animals of the same size and shape as predicted by Currie and Russell, but their 95% confidence intervals (CI) for many predictions are still enormous (which means, in plain terms, that the true proportions may have been very different to the predicted ones). For instance, Funston and colleagues scaled their Albertosaurus embryo dentary to a body length 715 mm, but with a CI range of 496–897 mm. Their (seemingly) larger Daspletosaurus specimen, represented by a toe claw, scaled to 1101 mm long, with a CI of 250–5954(!) mm. Estimates for their femoral lengths were also not well constrained, at least in Daspletosaurus, with CI values of 76–256 mm. Some of the upper ranges of these CIs are consistent with larger tyrant specimens that are already several years old, and they're thus far from the measurements we'd expect from embryos or hatchlings (Funston et al. 2020). Our most charitable assessment is that we're consistently getting the same basic signals about tyrant hatchling form, so we're probably on the right track, but we have a long way to go before we'll have reliable body proportion predictions for tiny tyrants. The discovery of more small tyrannosaurids would go a long way to helping this but, of course, a skeleton of an embryo or hatchling would be the best and most direct way to resolve any uncertainty.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Currie, P. J. (2003). Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the upper Cretaceous of North America and Asia. Canadian Journal of Earth Sciences, 40(4), 651-665.
Funston, G. F., Powers, M. J., Whitebone, S. A., Brusatte, S. L., Scannella, J. B., Horner, J. R., & Currie, P. J. (2021). Baby tyrannosaurid bones and teeth from the Late Cretaceous of western North America1. Canadian Journal of Earth Sciences, 58(9), 756-777.
Russell, D. A. (1970). Tyrannosaurs from the Late Cretaceous of western Canada. National Museum of Natural Sciences, Publications, in Paleontology, 1, 1-34.
Schmidt-Nielsen, K., & Knut, S. N. (1984). Scaling: why is animal size so important?. Cambridge university press.
Yes, against my better judgement, we're going there. (Publicity image for Jurassic World: Fallen Kingdom. This image is too populous online to find its original source, so let's just assume it's the PR department at Universal/Legendary Pictures.)
So, that’s that. With the release of Jurassic World: Dominion the so-called “Jurassic Era” — which is what certain posters now want us to call the six loosely connected films in the Jurassic Park series — is over. Whether it's actually concluded will surely be determined by box office revenues more than creative necessity but, whatever: for the time being, the Jurassic Park movie series is officially finished.
However we feel about the Jurassic films, we have to acknowledge two facts about them. First, they represent a uniquely successful stream of palaeontology-inspired products. I can’t think of another string of dinosaur movies that have all had theatrical releases, nor has anything dinosaur-related ever generated so many billions of dollars. Once enough time has passed to fully gauge their impact, I’m sure palaeontological historians will give the Jurassic films serious study as a cultural phenomenon that shaped decades of conversations about prehistoric life. Whether we like it or not, 21st Century dinosaur outreach takes place in a big, Jurassic Park-shaped footprint stamped into pop culture.
Second, it’s not controversial to say that the Jurassic series has been critically divisive. Only the 1993 original is regarded as a classic and is widely, deservedly considered to rank alongside Spielberg’s best crowd-pleasers like Jaws and Raiders of the Lost Ark. The Jurassic sequels, on the other hand, have made a lot of money, but fans and critics often clash about which, if any, rank above mediocre. 2015’s Jurassic World is generally regarded as the best sequel, perhaps aided by borrowing much of its plot structure from the original film, but even this has not escaped accusation of thin, contrived plotting and flat, boring characters. This is to say nothing of the series’ slide away from palaeontological science towards increasingly inaccurate, toyetic creature designs.
Moreover, and echoing broader trends in blockbuster cinema, the Jurassic films have also become increasingly action-orientated. This means, relative to the original, they feature many more dinosaur sequences. The new trilogy in particular is stuffed with as many dinosaurs as each film can bear. Box office receipts show that this elevated level of prehistoric mayhem has paid off, as least among general audiences and we can't truly blame the Jurassic filmmakers for adding more dinosaurs: they are, after all, making dinosaur movies. Aren't they just giving us what we want and expect? Maybe, but I suspect this is actually the fault line along which these films divide opinion. If you're the sort of person who punches the air every time a Jurassic film includes a new species, no matter how fleetingly and inconsequentially, you've probably enjoyed the last three films. If, however, you tire quickly of what can be repetitive dinosaur sequences and want a little more in terms of story and characterisation from your Jurassic experience, you're more likely to view this dino-centricity as mindless, dull prehistoric noise.
This raises the question of whether dinosaur films can, perhaps against expectation, go too far with their main draw: can they a dinosaur film actually have too many dinosaurs? The answer, of course, is a matter of opinion, but one way we might try to answer it objectively lies in revisiting the only Jurassic film we all agree is genuinely good: the original Jurassic Park. Were these filmmakers all in on dinosaurs, adding as many as their budget and technology would allow, or is the famously low dinosaur screen time of Jurassic Park a creative decision?
Now eventually you plan to have dinosaurs in your dinosaur film, right? Hello? Hello? Yes?
The story behind the script for the first Jurassic film is recounted in Shay and Duncan’s 1993 book The Making of Jurassic Park, and much of the following is taken from that source. The script took a long time to come together, going through several rewrites by different people. Original book author Michael Crichton was contracted to take the first stab at the film's screenplay but admitted that his heart was never in it. Crichton had literally just finished the novel and simply wasn’t interested in adapting the story so quickly after putting his own version to bed. A second treatment was penned by Malia Scotch Marmo, who’d just written Spielberg’s 1991 Peter Pan adventure Hook. Her version is notable for blending the character of Ian Malcolm with that of Alan Grant to give the latter more personality, as the weakly fleshed-out characters of Crichton’s novels were regarded as a problem that needed solving for the film.
Photograph of the essentially completed, but never used baby Triceratops built for Jurassic Park. The scene with this animatronic ended up being abandoned for creative reasons, despite the money invested in bringing it to completion. Fans would briefly see this guy in action during a very quick cameo in The Lost World, however. Image from Mike Tharme's Twitter feed.
But Marmo’s interpretation wasn’t well received either and, relatively close to the start of filming, another writer was hired for a third stab at cracking the story. Enter David Koepp, who you’re surely familiar with from some of the biggest blockbusters of the 1990s and 2000s: Jurassic Park and its first sequel, 1996’s Mission: Impossible, Sam Raimi’s Spider-Man, the 2005 War of the Worlds and… er… Indiana Jones and the Kingdom of the Crystal Skull (hey, I didn’t say they were necessarily good blockbusters). Koepp proved to be the person who could finally tap the full potential of Crichton’s novel, perhaps because he and Spielberg agreed on a major problem with their source material: it had too many dinosaurs. Lest it be thought I’m generalising or paraphrasing, this is exactly how Spielberg described adapting the novel.
“Believe it or not… the first thing I thought was that the book had too many dinosaurs in it. I didn’t think it was physically possible to make a movie that chock-full of dinosaurs… What I wanted to do was boil the book down and choose my seven or eight favourite scenes and base the script around those. So we crunched the book.”
Steven Spielberg, quoted in Shay and Duncan (1993, p. 12)
Given that the first Jurassic Park was pioneering so many new special effects, we might assume that Spielberg’s reservations reflect limitations of technology and budget. But while these were surely limiting factors, they were not the only considerations when it came to losing dinosaur sequences. In fact, we know expensive effects were scrapped after script changes in at least one instance, when a year’s worth of development and production on an animatronic baby Triceratops was abandoned late in pre-production. This effect was intended for a whimsical scene where Lex Murphy would ride around on it, further demonstrating the "dinosaurs were not monsters" ethos etched into the writing and production philosophy of Jurassic Park. But Koepp found the scene interrupted the flow and tone of the film wherever he placed it in the script. As he explained:
“If we put the [Triceratops] ride before the T. rex attack, it slowed down the movie; if we put it after the T. rex attack, why would this kid who has just been attacked by a giant lizard go and ride one?”
David Koepp, quoted in Shay and Duncan (1993, p. 64)
Because Koepp was working on the screenplay at the 11th hour, this decision meant that work on the 1.5 m long baby Triceratops robot was abandoned literally days away from its completion, all so that the film would have a tighter, leaner story. As much as Koepp felt that the audience needed a reminder that dinosaurs were “innocent” animals following the T. rex attack, a child riding a bounding baby Triceratops would have been a tonal shift too far, and certainly out of character for a traumatised child. With this dinosaur scene cut, Koepp added a little more whimsy to the foraging Brachiosaurus sequence, allowing our shell-shocked characters to be reminded that dinosaurs aren't evil or vindictive. They're just animals, as Grant puts it, doing what they do.
Concept art for the Jurassic Park rafting sequence, swiped from the Jurassic Wiki. This doubtless would have been an action-packed scene, but would it have added anything to the film?
This was not the only planned dinosaur sequence that was cut for pacing and tone. Several parts of the novel that seemed tailor-made for cinema were abandoned, such as Muldoon tranquillising the Tyrannosaurus and the famous river sequence. The latter, where Grant and the kids escape a swimming T. rex in a raft, was included in all previous drafts of the screenplay and went as far as having concept art produced, but Koepp removed it without hesitation. In discussing why, he addresses the “too many dinosaurs” issue directly, noting that Crichton's novel was actually bogged down by its large number of dinosaur episodes. He remarked that “It seemed to me that at certain points in the book we were being taken on sort of an obligatory tour past every dinosaur the park had to offer", such that "the raft trip was rather redundant” (Shay and Duncan 1993, p. 55). Clearly, Koepp didn't consider dinosaurs for the sake of dinosaurs an excuse for their inclusion in Jurassic Park: they had to add something to the film to justify their presence. The result is that dinosaur scenes only constitute about 15 minutes of Jurassic Park’s two-hour runtime. Their off-screen presence drives the film, of course, but almost 90% of the film passes without a dinosaur in shot.
If we turn to literature, we find that this kind of reserved approach to creating a dinosaur story is in good company. Other classic works of "dinosaur" fiction such as Jules Verne’s 1864 Journey to the Centre of the Earth, Conan Doyle’s 1912 The Lost World and Ray Bradbury’s 1955 A Sound of Thunder feature prehistoric animals in memorable sequences but, like the first Jurassic Park movie, these are kept short and impactful. I wonder if this reflects a shared realisation about the narrative potential of fictional prehistoric animals: as initially exciting as they are, they quickly exhaust what they can contribute to a story. Most fictional dinosaurs invariably have to interact with people and their roles are essentially limited to inspiring awe or terror, which means their actions are either peaceful or violent. We can vary where and why these interactions occur, and we may gain additional mileage from featuring different prehistoric species, but it’s difficult not to basically rehash the same ideas and scenes over and over once dinosaurs turn up in a story. And because dinosaurs are real animals, we can’t ascribe crazy, unexpected biology or properties to them, either — not with a straight face, anyway. Jurassic World; Dominion was never going to end the series with a reveal that an evil interdimensional dinosaur was the real villain of the series all along, or show that the dinosaurs were really birthed by an awful, vengeful Megadinosaurus queen. More the pity, perhaps?
My take on the most famous scene from Ray Bradbury's 1955 A Sound of Thunder. This short story is possibly the ultimate example of lean dinosaur storytelling as it features only a single, short interaction with Tyrannosaurus. Despite this, it remains a highly effective, thought-provoking tale and contains some of the best prose describing the appearance and movement of a large theropod ever written. Check it out if you haven't.
Compounding these creative issues for dinosaur films are the huge budgets needed to create dinosaur visual effects. Movie dinosaurs are so costly that their films must appeal to broad, mainstream audiences to be financially viable, and this means avoiding creative choices that will alienate casual viewers, especially children and families. Anything too scientific and “boring” is unlikely to feature, as is anything too violent or horrifying. This, I suspect, is why the Jurassic films are the only game in town for dinosaur motion pictures. Whether humans meet dinosaurs through time travel, in a “lost world” setting or via resurrection from fossils, the Jurassic films are already exploring the full remit of what movie dinosaurs can do. There’s probably not enough creative space for another franchise to present their own, fully distinguished take on dinosaur scenarios, especially given the potential financial losses if such efforts flopped.
Back to the Park
In representing its own contained franchise, the Jurassic series represents a unique case study of attempts to escape these creative restrictions. But it seems fair to say that, even after six instalments, the franchise never really figured out how to get more agency from its reptile stars. Repetition, not innovation, is the order of the day, leading to essentially the same moments playing out in each instalment (as super-seriously scientifically documented by Dave Hone, see his tweet below). Instead of new dinosaur dramas, we simply get more of the same dinosaur scenes. This suggests a creative ethos of, when in doubt, add more dinosaurs!
Got around to seeing Dominion last night thus allowing me to complete my table of Jurassic Park franchise cliches. There were, inevitably, loads. pic.twitter.com/NxvJKm9MLT
Ramping up the dinosaur content began in the first Jurassic sequels but reached its apogee within the most recent films. Whereas Jurassic Park slowly led us into the world of recreated dinosaurs and established its setting, characters and story before letting dinosaur havoc commence at the one hour mark, the sequels have started their action sequences earlier and earlier. Jurassic World has Velociraptors attacking their handlers after 25 minutes, people are visibly chomped in the first five minutes of Fallen Kingdom, and Dominion is the least patient of all, showing prehistoric animals attacking people within the first minute. And this is where we can start to explore whether an “add more dinosaurs” approach has drawbacks, because all these extra dinosaur scenes absorb time from the fabrics that actually tie films together: stories, characters and themes. And, OK, we might ask who is really watching a dinosaur film for great characters and stories, but we've seen that perfectly serviceable, universally-liked dinosaur films can be done (Jurassic Park) and, moreover, screenwriters should be aiming to have something to hang a film on to give their dinosaur action agency. Putting characters in peril is toothless if we don’t care whether they survive or not. Writing a "dinosaur film" doesn't excuse filmmakers from attempting to make the best, most engaging film they can.
Viewed from this perspective, Fallen Kingdom and Dominion are especially full of what is ultimately pointless dinosaur action, wheeling out prehistoric animals to menace our heroes for a short time (often less than a minute) before moving on to the next, equally pointless encounter. Portions of these films are like riding a dinosaur-themed ghost train where dinosaurs pop out to roar at us before disappearing into the shadows, never to be seen again. And lest it be thought I'm some sort of film snob (I'm not: my benchmark for enjoying most films is how closely they approximate Evil Dead II), the tedium of such scenes was not lost on Steven Spielberg himself, who has candidly spoken of how bored he was making the (relatively) dinosaur-heavy The Lost World:
“I beat myself up…growing more and more impatient with myself… It made me wistful about doing a talking picture because sometimes I got the feeling I was just making this big silent-roar movie… I found myself saying ‘is that all there is? It’s not enough for me.’”
Steven Spielberg, quoted in McBride (2011, p. 455)
It’s hard not to contrast this creative approach and the impact of these dinosaur sequences with those of the first film. Jurassic Park clearly relied on the tried and true creative philosophy of “less is more” and carefully managed its story and tone so that its dinosaur action scenes, once unleashed, were genuinely exciting. But the successive films seem to have embraced a “more is more” approach that prioritises dinosaur violence over anything else. For me, this is one of the main reasons that the Jurassic World series has been such a flatline: the overabundance of dinosaurs roaring and fighting starts to get in the way of the films, actually undermining my enjoyment despite, in theory, adding to the excitement. To give an example, here are six minutes of the Fallen Kingdom volcanic eruption set piece. This takes place, for context, about 35-40 minutes in:
What stands out here is, first; wow, there are heaps of dinosaurs packed into this segment, but second, most are disposable, throwaway additions. The Baryonyx, which is never named, set up or returned to, comes and goes within 90 seconds. Likewise, the Allosaurus menacing the tumbling gyrosphere is there for a moment, and then gone. Clearly, the most important dinosaurs are the stampeding collective: they're what is really driving the action and story at this point, along with the erupting volcano. To give credit where it's due, I actually find the fleeing dinosaurs and eruption pretty engaging (as ridiculous as the galloping ankylosaurs and so are ) and, as our heroes shelter behind a fallen tree being smashed to pieces by charging dinosaurs, I'm curious to see what happens next. But then everything stops... so we can have a quick dinosaur fight scene. No more eruption, no more stampede. All the scene's momentum is discarded so Carnotaurus can slowly stalk around the human characters before getting into a fight with Sinoceratops. Why these dinosaurs are fighting rather than fleeing like all the rest isn't clear, but within seconds none of it matters: the Carnotaurus is put down (killed? I'm not sure) by the passing Tyrannosaurus, which then stops to roar in defiance despite the island literally exploding behind it. The T. rex then leaves, paying no attention to the humans, and the eruption and dinosaur stampede resumes as if someone has thrown a switch offscreen.
Along with killing all momentum, this dinosaur fight is confusing and, uh oh, gets us asking questions about the film. Why weren't these dinosaurs running away? Why didn't the Carnotaurus predate the easily-caught people instead of breaking off to attack a multi-tonne horned dinosaur? Why did the T. rex attack the Carnotaurus and then just walk off? Does T. rex watch Parks and Recreation and wanted to help out Andy Dwyer? Where did all the stampeding dinosaurs go? While the Baryonyx and Allosaurus portions are so superficial and inconsequential that they don't hurt the flow of the film by themselves (if, admittedly, such incessant "dinosaur cameos" of the World films do become repetitive and grating, especially in Dominion), the Carnotaurus sequence totally distracts from what should be our main focus at this part of the film. I guess the logic was that giant reptiles fighting is exciting and will thus make the eruption more engaging, but it actually does the opposite: it sucks energy and drive from the movie. It also presents a problem for story progression because it leaves the film struggling to raise the stakes later on. How do you create a situation more dangerous and exciting than dinosaurs fighting on an exploding island? Doesn’t everything seem a bit flat and dull after that? These additions are so out of place that I strongly suspect they were only added because the film otherwise lacked large theropods fighting, as if that's the only way to put drama into a dinosaur film.
Unrelated clip from Jurassic Park.
The irony in this, of course, is that the World films also revel in nostalgia for the first Jurassic Park, and yet the creative philosophy behind them is almost antithetical to that used by Koepp and Spielberg. In my view, this it shows little understanding of what made the first film great. We talk a lot about how the revolutionary dinosaur effects of Jurassic Park were integral to its success, and how its portrayal of post-Dinosaur Renaissance science blew audiences away. We also acknowledge that Jurassic Park is a rare "lightning in a bottle"-style production, the output of some of the best filmmakers of the early 1990s working at the top of their game. These are all true points, but we should add “creative restraint” to this list of success factors. A “less is more” dinosaur philosophy allowed for a logical, well-paced story with likeable, charismatic characters and truly iconic, memorable dinosaur scenes. It might only be 12% dinosaurs, but that's enough to make their screen time special and satisfying without any risk of it becoming boring or repetitive. This allows the dinosaur sequences to be the emotional high points of the movie, all choreographed perfectly to the developing story. The super-tense mid-movie T. rex attack initiates the start of chaos on the island. The Velociraptor kitchen scene ups the ante as we approach the climax, suddenly throwing the kids — hitherto shielded by adults against danger — against two smart, deadly predators. And the climactic battle between Velociraptor and Tyrannosaurus finishes the film with a flourish, saving our heroes at the last moment in the most exciting way possible. These are dinosaur action sequences that build upon one another and drive the story, such that we know, intuitively, where we are in the movie. You needn’t look at your watch to know that the sight of the Tyrannosaurus roaring as the “when dinosaurs ruled the Earth” banner slips past means the film is over. This is blockbuster entertainment done with real craft and care, and it remains the best dinosaur film ever made not despite its lack of dinosaurs, but because of its lack of dinosaurs. It’s no surprise that the World films mine the iconography of Jurassic Park so frequently because, in never taking a break from throwing dinosaurs around, they never established compelling enough stories, characters or moments to create their own iconic elements.
So perhaps, as contradictory as it seems, the Jurassic film series makes a case that dinosaur films can work a lot better when they have fewer dinosaurs or, at least, when dinosaur action isn't prioritised over more fundamental and important components of filmmaking. The problems outlined here are not unique to the Jurassic sequels, of course: we could level the "too many dinosaurs" criticism at plenty of other films, from Peter Jackson’s 2005 King Kong to even the likes of 1966’s One Million Years BC. And I think it's important to add that "too many dinosaurs" doesn't necessarily ruin a film, but they might diminish our enjoyment to greater or lesser extent.
Ultimately the point made here is just a dinosaur-specific reminder that special effects and action alone do not make good films: it's memorable stories, characters and situations that resonate most with critics and audiences alike. So if this really is the end of the Jurassic franchise, let’s hope that the next generation of dinosaur films doesn’t just bring fresh ideas, fresh stories, and fresh palaeontological science to our screens, but that they also reflect on a crucial question for this niche genre. Can dinosaur movies have too many dinosaurs? Well, if you want everyone to enjoy your film, and you want to make a film that will last the ages, then maybe yes, yes they can.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
McBride, J. (2011). Steven Spielberg: A Biography, 2nd edition. University Press of Mississippi.
Shay, D., & Duncan, J. (1993). The Making of Jurassic Park. Ballantine Books.
Giganotosaurus carolinii carries the remains of a juvenile rebacchisaurid while sporting a bunch of crazy tissues on its face. Big scales, horns, and is that some kind of thick pad over its snout? Is all this artistic speculation or something inferred from fossils? Read on...
Up until now, my palaeoart career has not crossed paths with carcharodontosaurids, the gigantic, charismatic and famous allosauroids best known for Acrocanthosaurus, Giganotosaurus, Carcharodontosaurus and, most recently, Meraxes. This month, however, I finally had cause to restore Giganotosaurus carolinii, the largest of the group and, potentially, the largest of all theropods.
Carcharodontosaurids are, at first glance, not too challenging to restore: take an Allosaurus, turn everything up to 11 and job done, right? Well, maybe not. Not only are the proportions of carcharodontosaurids (and, to be fair, carcharodontosaurians in general) subtly different from their allosauroid ancestors, but their jaws and eye regions are characterised by a suite of complex sculpting and rugosities. It’s thought that these are epidermal correlates (Sereno and Brusatte 2008): distinctive bone surfaces and histological patterns that record different skin types interacting with the underlying bone (Hieronymus et al. 2009). I’ve written quite a lot about epidermal correlates at this blog because they provide heaps of important external soft-tissue information without direct soft-tissue fossilisation and learning to spot them, in my view, is an essential skill for any budding palaeoartist.
The presence of epidermal correlates on carcharodontosaurid skulls means that we can’t take an “anything goes” approach to restoring Giganotosaurus or its close relatives; instead, there probably is a “right”, or at least "more defensible", way to approach depictions of their faces. Alas, to my knowledge, no specific investigation has been conducted into what carcharodontosaurid skull textures represent despite our interest in other dinosaur epidermal correlates (e.g. Hieronymus et al. 2009; Carr et al. 2017; Delcourt 2018). This means there’s not yet a “go-to” study to provide artists with answers for restoring these animals and anyone wanting to illustrate Giganotosaurus credibly has to make their own interpretations from descriptions and illustrations in scientific literature. Having just been through this process myself, and realising that Giganotosaurus is a fan-favourite, I thought it might be of interest to share my thoughts here. I want to be upfront by declaring that the following deductions are little more than best guesses; without having direct experience of Giganotosaurus fossils I can’t write anything definitive about what Giganotosaurus looked like. Think of the following more as a discussion piece than a rigorous guide, and I welcome input and insight from others if I’ve made errors.
As I understand it, most of what's been illustrated of Giganotosaurus is shown above in this compilation of figures from Coria and Salgado (1995; greyscale graphics) and Novas et al. (2013; colour). Not as much as you might expect, right? There's a lot more known of this species that hasn't been published yet, some of which contain crucial information for palaeoartists (and I guess for scientists too).
And it’s in this spirit that, right off the bat, we need to mention that researching Giganotosaurus is pretty challenging. Its fossils are thinly documented despite Giganotosaurus being one of the more completely known carcharodontosaurids and, even today, almost 30 years since it was announced to the world, we only have a fraction more information available to us than when it was first named in 1995. Just a handful of its bones have been figured so good photos or illustrations of several fossils relevant to this conversation have not been published (Coria and Salgado 1995; Coria & Currie 2002; Novas et al. 2013). Thus, anyone trying to restore this animal from scientific papers alone will struggle for information and a lot of secondary sources — online photographs of fossils and casts, skeletal reconstructions and museum mounts etc. — are essential to obtaining basic information about its proportions and size, even if they risk introducing reconstruction errors. Closely related taxa and comparative descriptions (i.e. “Mapusaurus has a more rugose snout than Giganotosaurus”) are critical too, providing crucial details not mentioned in dedicated Giganotosaurus papers. I mention this because it means that, from the get-go, we’re not in an ideal research scenario for a palaeoartwork, and this makes the possibility of errors in interpretation all the greater.
With appropriate caveats established, let’s dive into this discussion. As with most theropods that have rugose, textured faces, our attention here is going to be on the bones of the snout and orbital region, as these are the principal areas to bear features that might signify epidermal tissues. The carcharodontosaurid fossil record contains a large number of maxilla bones (the main tooth-bearing bone of the upper jaw) and this is good news for artists, as the lateral surfaces of these potentially tell us a lot about the skin on the side of the upper jaw. The typical maxillary rugosity for carcharodontosaurids is well documented across several species, especially Carcharodontosaurus saharicus, Eocarcharia, Mapusaurus and Meraxes. It comprises a series of sub-vertical grooves and pits (Stromer 1936; Sereno et al. 1996; Coria and Currie 2006; Brusatte and Sereno 2007; Sereno and Brusatte 2008; Canale et al. 2022) and some species (e.g. C. saharicus, Eocarcharia, Mapusaurus) supplement these with prominent ridges extending along the base of the antorbital region. These bars separate the rugose maxillary body from the smoother bone of the antorbital fossa: that slightly impressed region of bone surrounding the antorbital fenestra.
The maxilla of Eocarcharia dinops, as illustrated by Brusatte and Sereno (2008), shows the texturing typical of carcharodontosaurid snouts. Note the ridge dividing the textured region from the smoother antorbital fossa: this feature isn't seen in all carcharodontosaurids but might tell us something about skin types all the same. The fossa region is particularly big in this species.
The texturing characterising carcharodontosaurid maxillae may be somewhat less pronounced in Giganotosaurus and thus, perhaps like Acrocanthosaurus, its maxillae may have been on the smoother end of the rugosity scale (Coria and Currie 2006; Eddy and Clarke 2011; Novas et al. 2013). All else being equal, this might imply differences in facial anatomy within Carcharodontosauridae: whatever those grooves and pits signify may not have been as exaggerated in some species as others. A caveat here is that, as is often the case with skin-altered bones, larger carcharodontosaurid individuals tend to have more exaggerated rugosity profiles than smaller ones (Coria and Currie 2006; Canale et al. 2014), suggesting a link with body size or age as well as differences between species. We probably want a number of maxillae from a range of differently aged individuals to establish whether a species has consistently smoother jawbones than its relatives.
Comparing these maxillary features with existing interpretations of dinosaur epidermal correlates provides potential insights into their significance. The textures in question are often likened to those adorning the lateral surfaces of abelisaur skulls and, if so, we might follow Delcourt (2018) in inferring that they represent scale correlates. This seems sensible to me, certainly more than other possible jaw coverings. Carcharodontosaurid maxillae lack the projecting rugosities consistent with armoured dermis or the branching neurovascular channels and oblique foramina found under beak or horn tissues (Hieronymus et al. 2009). Furthermore, the ridges bordering the antorbital regions in some carcharodontosaurids are inconsistent with beaks: cornified sheath tissues tend to terminate with obvious steps downwards into smoother neighbouring bone, not upwards to ridges of rugose bone (Hieronymus et al. 2009).
The skull of a common snapping turtle Chelydra serpentina. The more rugose parts of this skull correspond to regions covered in large scales, while the slightly finer rugosity around the jaws demark the distribution of the beak (also note the stepped topography at the beak/scale transition). Prominent ridges occur around the eyes and nose where large scales meet softer tissues: perhaps this is analogous to what we're seeing in those ridged carcharodontosaurid maxillae?
Raised bony ridges are seen, however, around the skull openings of reptiles with tough, tightly-adhering facial skin like crocodylians and certain turtles, marking some boundaries between thick, relatively immobile skin and softer, more flexible regions. We might expect the antorbital skin of theropods to flex slightly during breathing, as it does in birds, and I wonder if we're picking up some evidence of that in carcharodontosaurids? The notion that carcharodontosaurid maxillary skin might be tough and immobile is not without precedent, as early members of the broader Carcharodontosauria clade are thought to have had maxillary skin of this nature (Barker et al. 2019). If there really was a distinction in skin flexibility in the snouts of these animals it may have been obvious in life, as it is in crocodylians and turtles (I realise this sounds like advocating some form of shrinkwrapping— lightning flashes in the distance, thunder rumbles — but we can't overlook the fact that osteological features do, sometimes, correlate with skin types in living animals). I took these reptiles as inspiration in my reconstruction, giving Giganotosaurus a series of large, thick scales over the side of its upper jaw that terminate sharply around the antorbital region. I retained a full set of lips for reasons that have been thrashed out too many times to bear repeating here, except to mention that — like those of tyrannosaurs— carcharodontosaurid maxillae seem to constrain their rugosity to regions above the toothrow, suggesting whatever skin anchored above the labial foramina (the row of perforations along the jaw) was not so tightly anchored next to the teeth.
Immediately above the maxillae are another set of sculpted bones: the nasals and lacrimals. Collectively, these bones form the various fins and crests that line the top of the snouts in many allosauroids (see Chure and Loewen 2020 for a great visual of these), but the carcharodontosaurid condition is not typical of this wider clade. Nasal material is known for Giganotosaurus but it was not featured in its original description, nor (to my knowledge) has it been illustrated elsewhere. What’s hinted at in various reconstructions and papers is that Giganotosaurus joins Mapusaurus, Meraxes and Carcharodontosaurus in having especially sculpted nasal bones over the maxillary region, specifically bearing deep, generally parallel-sided grooves crossing transversely over the dorsal surface and vertically on the lateral face (Coria and Currie 2006).
The right nasal of Mapusaurus rosae, one of the better-illustrated examples of the crazy rugosities developed on these bones by some carcharodontosaurids. A shows the lateral view, B is dorsal, from Coria and Currie (2006).
These bones cannot be described as forming narrow crests as they can for Allosaurus and kin because their texturing meets in the middle of the skull and they are not pinched into long, narrow fins (Sereno et al. 1996; Coria and Currie 2006). Accordingly, some common artistic interpretations of these structures as supporting crests or a series of hornets over carcharodontosaurid faces (which I first assumed when embarking on this painting project, I think incorrectly: see below) may be erroneous: whatever skin made these features extended over the entire dorsal surface of the nasals as well as across the upper lateral region of the snout. Exactly what’s happening here is unusual among theropods, but the rugosity depth almost certainly implies some extensive cornificiation. I'll go further to say that, to the best of my knowledge, deep, subparallel grooves are uniquely associated with cornified pads growing at shallow angles to the underlying bone (Hieronymus et al. 2009). If correct, might we infer that heavy, thick bars of densely keratinised tissue adorned the top of carcharodontosaurid skulls? Cornified pads are predicted in this region elsewhere within Theropoda (e.g. within abelisaurids: Delcourt 2018) so such a suggestion isn’t entirely without precedent, but I'm not sure we've viewed carcharodontosaurids with such heavy ornament before. It would be great to see some actual research on this to investigate what’s really going on with these bones. Giganotosaurus striding around with fat cornified pads atop its face would be all sorts of awesome, especially given that we already think other regions of carcharodontosaurid faces might be adapted for headbutting (e.g. Sereno and Brusatte 2008; Cau et al. 2013).
An earlier version of my Giganotosaurus reconstruction with individual hornlets above the snout: thinking again on this topic, I probably got this wrong as the nasal texturing isn't consistent with the bones that underly hornlets in living species. I've already sent myself to bed without dinner as punishment.
The corrugated nasals are bordered posteriorly by further rugosities around the orbit. This is actually one of the better-known parts of the Giganotosaurus skull and it has been illustrated (Coria and Salgado 1995) so we can be pretty confident about what this region generally looked like, even if a lack of a comprehensive description means it’s difficult to know exactly what sort of rugosities it bears. In terms of basic structure, a rounded, horn-shaped process sits atop the lacrimal (the bone in front of the orbit) and a prominent boss projects above and somewhat laterally from the postorbital (the bone behind the eye). As seems typical for carcharodontosaurids and, indeed, for carcharodontosaurians in general, the latter slopes back and downward somewhat such that Giganotosaurus and kin probably looked perpetually worried, their postorbital bosses creating the appearance of a furrowed brow.
From what I can gather, the ultra-rugosity of the nasal bones doesn’t extend fully over the eyes in Giganotosaurus or its relatives. I suspect, based on what we see in better-illustrated carcharodontosaurids, that this reflects adornment of the lacrimal process with a cornified sheath rather than a pad. This creates the potential for a sharper horn than implied by the underlying bone shape, although it just as easily could be an exaggeration of the relatively blunt underlying bone structure. As is widely known in palaeoart circles, it can be difficult to predict the exact shapes cornified sheaths will take, even in modern species (Angst et al. 2020).
The postorbital boss variation of carcharodontosaurids, as illustrated by Sereno and Brusatte (2008): A shows the simpler morphology of Eocarcharia dinops; B shows Carcharodontosaurus saharicus.
Similar textures seem to have extended continuously onto the postorbital boss in derived carcharodontosaurids, such that we might imagine a continuation of the sheathed skin of the lacrimal onto this region (Coria and Currie 2006; Sereno and Brusatte 2008; Canale et al. 2022). There is, however, some variation of boss morphology within the clade in that some species have relatively smooth, rounded bosses (e.g. Sereno and Brusatte 2008; Cau et al. 2012): this is another area where more information specifically on Giganotosaurus would be welcome. For those species lacking pronounced texturing, I wonder if we’re dealing with big scale correlates rather than a surface covered with thick, densely keratinised tissue? These skin types may not be mutually exclusive however, as there is precedent for scale correlates showing signs of cornification in some dinosaurs (Hieronymus et al. 2009). A scaly postorbital boss in a young animal could well develop into a more cornified, horny structure in an adult. Again, more specimens of different growth stages might be needed here to be certain of true differences between species.
The result of all this noodling: Giganotosaurus looking a little more knobbly than usual, and also a bit world-weary thanks to that postorbital boss. Maybe the pressure of the "who's the biggest theropod" competition is pretty intense for these guys.
Putting all this together resulted in the image of Giganotosaurus that accompanies this post. Thanks especially to the big cornified pad bridging the middle of the skull, this is a face that looks more heavy-duty than we’re used to and maybe less generically “allosaurian”. But unfamiliar as it is, I’m happy with this outcome because following evidence to unexpected results is one of the great joys of palaeoart, and I always enjoy rationalising an unusual reconstruction from a foundation in science rather than mere speculation. But, again, I want to stress that this is just my interpretation of information gleaned from a less-than-ideal representation of Giganotosaurus in technical literature. This means I may have made errors obvious to those more experienced with these fossils and, moreover, when the structures discussed here are finally studied for their soft-tissue significance, the outcomes may be very different.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Barker, C. T., Naish, D., Newham, E., Katsamenis, O. L., & Dyke, G. (2017). Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii. Scientific Reports, 7(1), 1-8.
Brusatte, S. L., & Sereno, P. C. (2007). A new species of Carcharodontosaurus (Dinosauria: Theropoda) from the Cenomanian of Niger and a revision of the genus. Journal of Vertebrate Paleontology, 27(4), 902-916.
Calvo, J. O.and Coria, R. (1998). New specimen of Giganotosaurus carolinii (Coria & Salgado, 1995), supports it as the largest theropod ever found. Gaia, 15, 117-122.
Canale, J. I., Apesteguía, S., Gallina, P. A., Mitchell, J., Smith, N. D., Cullen, T. M., ... & Makovicky, P. J. (2022). New giant carnivorous dinosaur reveals convergent evolutionary trends in theropod arm reduction. Current Biology.
Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7(1), 1-11.
Cau, A., Dalla Vecchia, F. M., & Fabbri, M. (2012). Evidence of a new carcharodontosaurid from the Upper Cretaceous of Morocco. Acta Palaeontologica Polonica, 57(3), 661-665.
Coria, R. A., & Currie, P. J. (2003). The braincase of Giganotosaurus carolinii (Dinosauria: Theropoda) from the upper cretaceous of Argentina. Journal of Vertebrate Paleontology, 22(4), 802-811.
Chure, D. J., & Loewen, M. A. (2020). Cranial anatomy of Allosaurus jimmadseni, a new species from the lower part of the Morrison Formation (Upper Jurassic) of Western North America. PeerJ, 8, e7803.
Coria, R. A., & Currie, P. J. (2006). A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina. Geodiversitas, 28(1), 71-118.
Coria, R. A., & Salgado, L. (1995). A new giant carnivorous dinosaur from the Cretaceous of Patagonia. Nature, 377(6546), 224-226.
Delcourt, R. (2018). Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific reports, 8(1), 1-12.
Eddy, D. R., & Clarke, J. A. (2011). New information on the cranial anatomy of Acrocanthosaurus atokensis and its implications for the phylogeny of Allosauroidea (Dinosauria: Theropoda). PloS one, 6(3), e17932.
Hieronymus, T. L., Witmer, L. M., Tanke, D. H., & Currie, P. J. (2009). The facial integument of centrosaurine ceratopsids: morphological and histological correlates of novel skin structures. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology: Advances in Integrative Anatomy and Evolutionary Biology, 292(9), 1370-1396.
Novas, F. E., Agnolín, F. L., Ezcurra, M. D., Porfiri, J., & Canale, J. I. (2013). Evolution of the carnivorous dinosaurs during the Cretaceous: the evidence from Patagonia. Cretaceous Research, 45, 174-215.
Sereno, P. C., & Brusatte, S. L. (2008). Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger. Acta Palaeontologica Polonica, 53(1), 15-46.
Sereno, P. C., Dutheil, D. B., Iarochene, M., Larsson, H. C., Lyon, G. H., Magwene, P. M., ... & Wilson, J. A. (1996). Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. Science, 272(5264), 986-991.
Stromer, E. (1936). Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. VII. Baharije-Kessel und -Stufe mit deren Fauna und Flora. Eine ergänzende Zusammenfassung. Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Abteilung n. f., 33:1–102.
"Hi, I'm the 1/35 unpainted Beasts of the Mesozoic Tyrannosaurus. You may remember me from this Kickstarter campaign and the Creative Beast Studio website. But now I'm being reviewed at this blog to evaluate my box claim of being an 'ideal 3D reference for palaeoart'. How do I fare? Read on!"
Now here’s something I never thought I’d have cause to write: a review of a commercially available dinosaur model marketed, at least in part, as a palaeoart reference. It’s testament to the rising popularity of palaeoart and the growth of the number of practitioners that products are now being advertised specifically to people who don’t just enjoy palaeoart, but enjoy creating palaeoart.
The model in question is Creative Beast Studio’s new Beasts of the Mesozoic 1/35 unpainted Tyrannosaurus, a poseable dinosaur figure with the very words “ideal 3D reference for paleoart!” printed on the box. David Silva’s Beasts of the Mesozoic (hereafter BotM) line has been creating quite a stir among collectors for bringing the detail and articulation of modern action figures to dinosaurs. The current range includes dromaeosaurs and ceratopsians, and the tyrannosaur line is inbound. The frequency of some models being sold out from online stores is a testament to their popularity and, with individual price tags mostly upwards of $50, BotM products are clearly in the realm of poseable models for grown ups rather than children's action figures.
Despite not being much of a dinosaur model collector myself, I preordered the BotM Tyrannosaurus specifically because I saw its potential as an artistic reference. The preorder price was $65 + shipping, which is the same as the preorder cost for the fully painted version that will be released later this year. I received my order earlier this month and it’s joined a small collection of other bits and pieces used in my efforts at restoring ancient animals, including model animals, replica fossils, 3D printed materials and my own crude constructions of wire, cardboard and sculpting materials. I feel that building animals in 3D before painting them is probably the best way to approach palaeoart — it was, after all, the practise that Charles Knight swore by — but the time, space, money and skills needed to create 3D models for every restoration are not available to everyone. Digital models, either made ourselves or sourced online, avoid some of these issues but are arguably less satisfying and informative to work from than physical ones. Holding and manipulating a real object conveys information about form and proportions that we may not get when viewing a 3D representation on a screen, and I find it much easier to experiment with light and shadow in the real world. Don’t get me wrong: I’ll use a digital model over nothing, but I’d much rather have a real, physical reference than a virtual one if given a choice.
I’m thus fully onboard with the sale of quality palaeoart reference models and — to my knowledge —the BotM Tyrannosaurus enters this field unchallenged by competitors offering models specifically as palaeoart reference aids. Indeed, it's rare to see an artistic reference offered for any type of reptile, as animal art guides are terribly biased towards mammals and birds. In this sense, advertising the BotM Tyrannosaurus as an art reference is quite exceptional, but being the only player on the field doesn’t tell us anything about quality. Let's get into this: how does the BotM Tyrannosaurus fare as a reference tool, and what are you getting for your $65?
Straight out of the box, it’s apparent that the BotM Tyrannosaurus is a high-quality product. The moulding detail is superb and it doesn't look or feel at all cheap or plasticky. At around 35 cm long the model is big enough to view its details without it dominating your workspace, and this was the main reason I opted for the 1/35 version and not its gigantic, almost 70 cm long 1/18 counterpart (even if that does look extremely cool). Although possessing a satisfying heft, the model is small and light enough to hold comfortably in one hand, which is a big plus for something you’re potentially drawing from.
To show off the detailing and reference potential of the BotM Tyrannosaurus, I took it outside to my garden for these shots. The sculpting picks up light and shadow extremely well and looks great even on a phone camera. Note that, among other poses, you can achieve the Prehistoric Planet courting posture if you want the first-hand experience of being flirted with by an 8-tonne murder reptile.
Not that you need to hold this model for it to retain a pose. A variety of interchangeable lower leg components are provided that can be used to create a range of postures and demeanours. The two solid legs are probably your default setting as they provide a stable base for free-standing on even slightly uneven surfaces, and you can make plenty of adjustments to the head, torso and tail before the model topples over. If you’re after something more dynamic, you can swap in the mobile leg pieces which, in addition to adjustable ankle joints, permit attachment of different feet to create distinct parts of a step cycle. The solid legs can be pinned to a base so that, when paired with an adjustable leg, the model can be positioned as if running or walking (although it's not recommended to leave it in this state permanently). Across the body, some 22 points of articulation allow for posing the jaws, head, neck, mid-torso, tail, legs and arms, often offering rotation as well as extension and flexion. The tail is articulated in several places and sports a wire-infused end piece that can continue any arc made with the more proximal, jointed segments.
I initially found the joints to be extremely stiff to the point where I couldn’t distinguish their rigidity from having reached the figure’s arthrological limits. Paperwork provided with the model acknowledges this and recommends heating the joints in water or with a hairdryer rather than forcing them, and this indeed loosens them somewhat. Happily, early fears that I was going to have to get the hairdryer out every time I wanted to adjust the pose have not been borne out as joint mobility has improved substantially as I’ve worked the model through its paces, so far without any sign of compromising stability.
As you can see in accompanying photos, the model does a great job of filling space around its joints so that we retain that classic T. rex shape no matter what pose we choose. I’m particularly impressed with the adjustable hood-like neck piece that hides the articulation for the neck and head: once you’ve twisted and rotated that head to where you want it, the neck piece can be pulled into the right place. This retention of form around joints comes at the price of some mobility, however, and the posing options are really more about varying standing and walking poses than exploring the full repertoire of joint motion available to a real T. rex. From an art reference perspective, it would be neat to have more flexibility but, realistically, there must always be engineering compromise in a figure like this, especially if we’re also expecting it to stand on its own feet. In all, given the challenges of creating jointed dinosaur models, the BotM Tyrannosaurus is extremely well-executed and looks far better than anything else I've seen attempting the same goal. Even the mouth closes nicely with a very respectable oral seal, despite a wealth of complex internal detail.
Moving on, I quickly want to mention the colour, even if this seems an odd thing to bring up for an unpainted, entirely grey model. I don’t know how much thought went into the specific shade of grey but, in any case, the choice of medium-dark grey works well: it’s neither too light or dark to obscure shadows or highlights and it photographs well. As an aside, I also think this thing looks great as a grey, uniform figure perched on a desk or shelf: it somehow looks more timeless and informative than any of the painted models I have. The option exists to paint this model yourself, of course, but I have no plans to: it works much better as an art reference if it remains a blank canvas.
More images from Tyrannosaurus trip to the garden. I was going for the Denver Museum of Nature and Science's famous high-kicking T. rex on the right, but I think I achieved something closer to Tyrannosaurus after it's stepped on Lego.
Of course, all this fine production will be for nought if the reconstruction itself is off, so what about the anatomy? In short, the BotM Tyrannosaurus is an excellent T. rex restoration for 2022. We would expect nothing less given that it was sculpted by Jake Baardse, the digital sculptor behind the awesome Saurian T. rex (among other excellent artworks). The body proportions are well captured with the laterally expanded and blocky posterior skull region, barrel-shaped torso and deep pelvis that characterise adult Tyrannosaurus all present and correct. Modern soft-tissue highlights include lips, correctly placed nostrils and ear openings, as well as well-rendered muscles of respectable position and volume. In terms of soft-tissue bulk, it hits the sweet spot looking like a healthy animal, being neither too lean nor too tubby. All this comes together so that, unlike many (perhaps most?) Tyrannosaurus models, this undoubtedly represents T. rex rather than a generic carnivorous dinosaur, a generic tyrannosaurid or — shudder — a Jurassic Park knock off.
It would be remiss not to mention the enormous amount of fine details in the sculpt. They include tarsal scutes, an individually crafted tongue, skin creases, calluses, large facial scales and bosses above the eyes. Some of these are necessarily speculative — remember that, for all its fame, we’re still mostly in the dark on exactly what Tyrannosaurus looked like — but it all feels appropriately grounded in what we know of theropod dinosaur soft-tissues. There’s nothing here that anyone can firmly point to as “wrong”. I wonder if some will lament the absence of protofeathers, but the wholly-scaly approach is probably better for an art reference than one where body contours are buried under speculative tufts. What we’ve essentially got here is the foundation anatomy you must include when drawing Tyrannosaurus, over which artists can accessorise or augment based on their own views. Perhaps the only genuine anatomical quibble I have is that the extremely fine (mostly submillimetre) scales across the body are too big for a tyrannosaur at this scale (T. rex scales were tiny, just 1-2 mm across) but I can look past this. It gives the model an appropriately rough texture rather than a gleaming, smooth finish and, hey, if nitpicking scale size is the only real complaint with a dinosaur restoration, that’s normally a sign of a job well done.
With the BotM Tyrannosaurus acing every test, we’ve reached a final question: the above is all well and good, but does anyone need a Tyrannosaurus palaeoart reference figure, what with the near infinite numbers of specimen photos, diagrams, 3D scans, model skulls and skeletons etc. that are also available? Surely this is something that’s just “nice to have” rather than essential? To test this, I returned to one of my previous Tyrannosaurus restorations to see how it stood up to the BotM sculpt and… I immediately noticed errors in my work. Not major glaring super errors, but things that I’d want to get right and was glad to fix. Our familiarity with T. rex makes it easy to forget that Tyrannosaurus was a pretty unusual theropod and, while it’s easy to draw something that approximates it, completely nailing T. rex can be an artistic challenge even with a whole folder of 2D reference material. I’ve yet to craft a whole illustration using the BotM T. rex but I can already see it’s going to be a valuable palaeoart aid when that happens.
What difference does a good 3D reference make? Comparing an image I completed a few months back with the BotM Tyrannosaurus revealed a few goofs, some gleaned from less-accurate references, and others from my own miscalculations of body shape. The biggest adjustments here concern skull shape and leg position, both of which are much better in the right image than in the original. And yes, this dark, dark image is probably the worst example I could have used for showing the positive effects of this model on refining T. rex anatomy. Slow clap.
And I think that speaks for itself as a conclusion. Is the Beasts of the Mesozoic Tyrannosaurus ideal for crafting palaeoart, as it says on the box? Absolutely, and it’s a strong recommendation from me to grab one if you’re in the regular habit of drawing king tyrants. It may, indeed, be the single best artistic reference tool for drawing adult Tyrannosaurus available and we’d be absolutely fine if the world was flooded with T. rex artwork based on it. To that end, my fingers are crossed that other species in the BoTM range get similar art reference treatment, being scaled to handheld proportions and released with a flat grey colouring. A BotM line aimed at artists would not only be extremely useful for individual palaeoart practitioners but would benefit palaeoart as a whole: like all artforms of natural things, development and investment in quality reference resources can only help our collective understanding and ability to visualise our subject matter, and stronger, more interesting art is the result.
The Beasts of the Mesozoic 1/35 Tyrannosaurus is available to preorder from here, as is its bigger 1/18 counterpart and the upcoming range of tyrannosaur figures. The full Beasts of the Mesozoic line can be viewed at the Creative Beast Studio website.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
Predicting what will become a palaeoart meme is a dark, mysterious art. Sometimes news drops that should, given everything we know about the folks who create and like palaeoart, go absolutely viral. It should be illustrated again and again, find its way into books, magazines and maybe even documentaries, and inspire so much online content that old, miserable people like myself become quietly bored and tired of seeing it. But not all news of this sort takes hold among artists and, for whatever reason, it falls through the cracks.
Enter, stage left, the decade-old proposal that consumption of Triceratops carcasses by Tyrannosaurus involved the literal decapitation of the horned dinosaur corpse. Holy cow, how did we miss that one? Initially pitched in a conference poster at the 2012 SVP meeting by Denver Fowler and colleagues, the “How to eat a Triceratops” hypothesis has made a decent splash outside of the palaeoart community. It was featured in NatureandNew Scientist among many other news outlets back in 2012, and the then-active Walking with Dinosaurs website turned it into a short film. The Smithsonian mounted the Nation's T. rex specimen gripping a Triceratops frill, an action hypothesised by Fowler et al. as necessary to get at the neck steaks beneath. But even with widespread sharing of a Nate Carroll graphic operating as an instruction manual for palaeoartists (below), the internet has not been inundated with images of Tyrannosaurus ripping the heads of horned dinosaurs, aside from rare examples like Luis Rey's take. I can't be the only one finding this strange, especially given the amount of T. rex art out there. Come on, people: it’s T. rex pulling the head off Triceratops! Were we asleep in 2012? As you've already worked out, the image above is my atonement for missing such an awesome source of palaeoartistic inspiration.
Nate Carroll's guide to eating Triceratops necks, if you're a T. rex. A, grab frill; B, use the frill as a lever to tear the neck; C, pull the head off; D, eat. It's not quite as straightforward as ordering a pizza, but you can't argue with the results. (From SciTechDaily)
Of course, and as Denver has noted on his website in response to the press interest in this hypothesis, we need to tread carefully around the “How to eat a Triceratops” data because it hasn’t been peer-reviewed and published yet. A paper is on the way but, for now, what’s suggested in the abstract is exciting and compelling. A collection of c. 100 Triceratops was examined for bite marks to reveal a large number (maybe as high as c. 18%) with scores and punctures attributable to T. rex teeth. It’s rare to allocate theropod bites to a single species but, among the very latest Cretaceous deposits in western North America, Tyrannosaurus is the only animal that was capable of leaving gigantic punctures and gouges in dinosaur bones. And if that's not convincing enough, casts of tooth marks sometimes replicate T. rex dental morphologies with precision (Erickson and Olsen 1996). Using these criteria, dozens of hadrosaur and horned dinosaur specimens with bites from Tyrannosaurus, as well as some T. rex bones with cannibalistic feeding traces, have been identified in recent decades (e.g. Horner and Lessem 1993; Erickson and Olsen 1996; Carpenter 1998; Happ 2008; Longrich et al. 2010; Depalma et al. 2013; Mclain et al. 2018). This work is all so recent because historic collection and examination practises tended to overlook T. rex feeding traces, so we're only now learning how common — relatively speaking — these marks are.
Tyrannosaurus tooth marks on horned dinosaur frills have been reported outside of the Fowler et al. study, suggesting whatever behaviour these traces represent may have been widespread and routine. These examples are from Longrich et al. (2010): C is only tentatively identified as a ceratopsid frill element, but D is confidently identified as a right squmosal (i.e. the bone forming the right lateral frill region).
What specifically underpins the “How to eat Triceratops” hypothesis are bite marks in specific places on the back of Triceratops skulls. Specifically, multiple specimens show tooth gouges and punctures on Triceratops frills, and these are difficult to explain as actual feeding traces. So far as we can tell, there wasn’t much to eat on this part of the Triceratops body. Perhaps, instead, they represent carcass manipulation marks, where the head was adjusted and pulled about to move the corpse into a more accessible position? But there's more: Fowler et al. (2012) also report tooth traces on Triceratops occipital regions: parts of the skull situated deep within Triceratops neck tissues that would only be accessible on heads separated from their necks. It’s not much of a jump to link these traces: maybe all that jostling with the frills wasn't really about moving the whole carcass, but specifically to get at the neck soft-tissues? While the frill was probably an obstruction to biting the voluminous cervical musculature on a living Triceratops, in death it may have been a useful lever with which to manipulate and pull at the head. Given enough pulling, twisting and brute force, that mighty Triceratops head would eventually tear off: dinner is served, as they say.
Indirectly supporting this idea is good evidence that T. rex feeding could be very destructive in general, even when consuming animals as large as Triceratops. One of the most famous specimens to record Tyrannosaurus bite marks is a Triceratops pelvis described by Greg Erickson and Kenneth Olsen in 1996. Riddled with up to 80 tooth marks across several surfaces, this gigantic limb girdle was clearly moved around a lot by the feeding Tyrannosaurus (or tyrannosauruses) and chunks were literally shorn off by powerful bites, including one of the iliac blades and (almost) half a vertebra. The latter only remained attached by a small amount of bone and Erickson and Olsen ascribed this to the act of separating the pelvis from the rest of the body: a tremendous feat if it happened. Given this specimen, and the wealth of other fossils demonstrating the strength and force of a feeding Tyrannosaurus, I can totally buy that T. rex could decapitate Triceratops carcasses to access a bounty of horned dinosaur neck meat.
A caveat to all this, and a particularly necessary one in case we get swept along by the T. rex hype train, is that we shouldn’t imagine major dismantling of Triceratops carcasses taking place with freshly killed or otherwise untouched bodies. Neat as it is to imagine Tyrannosaurus ripping the head from a freshly-vanquished Triceratops, waving it aloft and roaring triumphantly like some kind of 8-tonne Predator, modern animals generally follow reliable carcass consumption patterns where easily accessed and nutritious tissues are eaten before difficult-to-access or less-nutritional parts (Blumenschine 1986). Typically, animal hindquarters are eaten first, then the contents of the abdominal cavity, followed by the forequarters and any fleshy bits on the skull, then the limb bones, and finally the internal contents of the head. Under this model, we might place Triceratops neck tissues as “mid-priority” fodder: decent enough eating to make them desirable, but only worth the energy and time investment of bypassing the head if more sought-after parts of a carcass are gone. I’ve attempted to show this in my artwork above by depicting the legs and arms of the Triceratops as already consumed, and the ribs are already exposed from the body being opened to eat the internal organs.
The image at the top of this post isn't my first dance with the Triceratops decapitation hypothesis. In this painting from earlier this year, the decapitated Javelina Formation ceratopsid is meant to be the result of tyrannosaur activity that preceded the arrival of more noble, elegant creatures who'll clear up the mess.
And that's where I'll leave things today. As noted above, a paper on all this is in the works and I'm looking forward to reading it when it comes out. I'm resisting the temptation to springboard onto other topics related to T. rex tooth marks: feeding habits, neck and jaw strength, and their embarrassment of older considerations of Tyrannosaurus tooth and jaw strength (“...its viscous-looking teeth were not as bad as they seemed: if it had tried to tackle living animals, the teeth would have snapped off in the struggle” - oh, Halstead 1975. that comment has not aged well). But time isn’t on my side and we’ll have to save that for another time. Or maybe we’ll finally move away from posts about big theropods. There is a good reason for all this, honest.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Blumenschine, R. J. (1986). Carcass consumption sequences and the archaeological distinction of scavenging and hunting. journal of Human Evolution, 15(8), 639-659.
Carpenter, K. (1998). Evidence of predatory behavior by carnivorous dinosaurs. Gaia, 15, 135-144.
DePalma, R. A., Burnham, D. A., Martin, L. D., Rothschild, B. M., & Larson, P. L. (2013). Physical evidence of predatory behavior in Tyrannosaurus rex. Proceedings of the National Academy of Sciences, 110(31), 12560-12564.
Erickson, G. M., & Olson, K. H. (1996). Bite marks attributable to Tyrannosaurus rex: preliminary description and implications. Journal of Vertebrate Paleontology, 16(1), 175-178.
Fowler, D.W., Scannella, J.B., Goodwin, M.G., & Horner, J.R. (2012) How to eat a Triceratops: large sample of toothmarks provides new insight into the feeding behavior of Tyrannosaurus. Journal of Vertebrate Paleontology 32(5, abstracts vol): 96
Halstead, L. B. (1975). The evolution and ecology of the dinosaurs. P. Lowe.
Happ, J. (2008). An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. In Larson P. & Carpenter, K. Tyrannosaurus rex the Tyrant king, Indiana University Press. p. 355-370.
Horner, J. R., & Lessem, D. (1993). The complete T. rex. Simon & Schuster.
Longrich, N. R., Horner, J. R., Erickson, G. M., & Currie, P. J. (2010). Cannibalism in Tyrannosaurus rex. PLoS One, 5(10), e13419.
Mclain, M. A., Nelsen, D., Snyder, K., Griffin, C. T., Siviero, B., Brand, L. R., & Chadwick, A. V. (2018). Tyrannosaur cannibalism: a case of a tooth-traced tyrannosaurid bone in the Lance Formation (Maastrichtian), Wyoming. Palaios, 33(4), 164-173.
Tabletop gaming and (scientifically credible) dinosaurs: together at last! The draft cover of Nathan Barling's Dr Dhronlin's Dictionary of Dinosaurs, a book illustrated with my palaeoart and now being crowdfunded over at Kickstarter.
Time to announce a new project that, I must admit, I never saw coming. For the last few months, I've been working with insect palaeontologist and taphonomy expert Nathan Barling to create a new book: Dr. Dhrolin's Dictionary of Dinosaurs: a palaeontologically-informed, palaeoart-heavy supplement for your tabletop roleplaying adventures. I'm aware that there are enough awesome keywords in that sentence to get some folks on board so, if you're already sold, head to the Kickstarter page for full details.
Still here? OK, here's some extra information and background. This project is, by far, Nathan's baby and I'm really only involved as an artist and paleontological consultant, along with fellow advisors David Hone (whoever he is) and pterosaur expert Michael O'Sullivan. Nathan, who I used to teach back in his undergrad days, approached me about illustrating this book at Christmas last year knowing full well that I don't do the whole tabletop gaming thing. My entire experience with such gaming was condensed into one evening about ten years ago, so everything I know about it comes from cultural osmosis. I believe it involves a traditional fantasy setting, dice, campaigns run from behind little cardboard houses and... Jeremy Irons? He's part of this somehow?
Fortunately for us all, Nathan wasn't interested in my knowledge of RPGs. Instead, he wanted my art so he could create a 5th Edition supplement featuring modern, scientifically-informed takes on prehistoric animals. There are, I understand, already some dinosaurs in official D&D canon, but they're apparently pretty "standard" and not especially accurate to their true palaeobiology. Realising that the reality of dinosaurs is way more interesting than their pop-culture stereotypes, Nathan wants to bring a diversity of extinct animals to your campaigns, each with stats and abilities inspired by their real anatomy and hypothesised behaviours. He's also taking inspiration from palaeoenvironmental reconstructions of specific geological formations to create new, science-informed worlds for your quests to take place in. Furthermore, he's creating palaeo-based player races that should be new and interesting, not predictable and familiar. With this supplement, you'll be able to play as folks inspired by azhdarchoids or obscure ornithischians rather than generic "dinosauroids". I'm sure we're going to meet all these goals. Even writing as I am — someone totally ignorant of this vast topic — I'm pretty confident that there aren't many palaeontology 5th Edition projects being guided by four published, PhDed scientists. If you've ever felt your tabletop campaigns were lacking a Yutyrannus ambush, a surprise encounter with Gigantspinosaurus or a Microraptor player companion, this is the book for you.
To bring all this to life, Nathan has full access to my artwork portfolio and is also commissioning me to do new pieces, both of species I've not yet painted as well as new works showing adventurers interacting with scientifically-credible extinct animals. You can get a flavour of what the latter will involve from the cover, which has already been painted and (provisionally) designed, below (NB: a professional designer will be putting everything together next year, so what you see here and at Kickstarter is only indicative of the final product, not finalised book content). This piece was very much a collaborative effort: I can handle dinosaur art well enough, but Nathan's got a tight grip on the more fantastical content and is steering me accordingly. I was thoroughly told off for including an orb staff in an earlier iteration of this image, which I now understand is the tabletop adventure equivalent of legwarmers.
Witton does 5th Edition art: two Utahraptor take on a band of travellers, including one of the new player races, the Pterochaps (OK, OK, actually called "the Children of Seth"). Can you spot all the palaeo references on the adventures? You're looking for azhdarchid pterosaur wings, a Tyrannosaurus skull (in anterior view), a juvenile Psittacosaurus skull and a bunch of ceratopsian-inspired costuming. No, you're a big dinosaur nerd.
The Kickstarter for DDDD (which, I confess, is not the most elegant acronym) went live yesterday morning and we've been totally blown away by the response. The £9000 minimum needed to get things moving was met by lunchtime and, at the time of writing (Sunday evening) the project has over £40,000 in pledges. Wow, and thanks to everyone who's pledged something already. Nathan's promotional efforts have really paid off (and he, indeed, deserves all the credit for this, I've done very little despite my name being on the draft cover). What this means is that DDDD is definitely happening and, if you want in at ground level, now's the time to sign up, especially if you want access to the Kickstater tiers with additional rewards. All being well, you'll be holding physical copies of Dr Dhronlin's Dictionary of Dinosaurs by late next year, which means we need to get to work. Here's that Kickstarter link again, and I'll see you on the flip-die. That's what you tabletop guys say, right? Because of the dice? Hello? Is this thing on?
The very first image posted at this blog, way, way back in November 2012. It shows a coloured version of a piece I created for the Pterosaur.Net blog earlier in the same year. How time — like so many quad-launching, cowboy-riding azhdarchids — has flown.
I don’t normally worry about blog anniversaries, but today marks a full ten years since I started writing this blog and a decade of writing and uploading artwork in the same venue feels like an achievement worth mentioning. According to Blogger stats, over 3 million people have checked in here in the last decade and, while I have no idea how genuine that number is, it implies someone is reading this stuff, even if it's just bots. If you are among those who have stopped by in the last few years, know that your visit is appreciated: I owe a big thanks to everyone who has read one of my posts, left a comment or shared my articles and artwork around the internet. And that applies to the bots, too: thanks for stopping by, fellas, and for all your weirdly-worded comments trying to get us to click stuff. We're not going to, but it's nice that you try.
It’s funny looking back on the very first days of this blog. I initially imagined this would be little more than a “picture of the day/week” style affair to promote my artwork in the wake of finishing my first book: Pterosaurs: Natural History, Evolution, Anatomy. That explains two pterosaur-themed first posts, and also why the third featured Tyrannosaurus: I chose T. rex as a palate cleanser after drawing and writing about pterosaurs for several years. There’s an ironic twist to this that I can’t reveal yet: let’s just say that the roles of those taxa might end up being reversed soon. A newly-released book called All Yesterdays formed the subject of post five, and my coverage of that saw the abandonment of any pretence that this would be a short-format blog.
From post 3, my first attempt to restore Tyrannosaurus as a semi-professional palaeoartist. It's pretty wonky to my modern eyes and — shock — even has exposed teeth. That won't fly in a few years, 2012 Mark, and we might want to talk about that facial reconstruction too (and, hey, 2022 Mark, when will you finish writing the paper on that?). This image would resurface a few years later in a modified form for one of the most popular articles on this site: "Revenge of the scaly Tyrannosaurus".
Since then, we’ve covered a fairly broad set of topics within palaeontology, with subject matter mostly divided across the science of extinct animal life appearance, deep-dives into animal palaeobiology, portrayals of palaeontology in the media, and the history of palaeoart. Although I’ve enjoyed writing virtually everything I’ve posted, a few pieces stand out as personal favourites. They aren’t the most popular articles, but those posts where I sought to answer a simple question and found a complex rabbit hole to explore, or the ones where I found conventional wisdom was incorrect and I could present an alternative based on peer-reviewed publications. Among these are my three articles on geomythology (covering the alleged fossil origins of griffins, dragons, cyclopes, unicorns and others), all of which found reason to question mainstream views linking certain fossils and mythological creations. I also enjoyed digging into the literature on the many fraudulent claims about mastodon hair: who’d have thought, with thousands of images of mastodons covered in brown, shaggy hair, that our evidence for such an integument would be near zero? In truth, writing about anything where the mainstream interpretation is at odds with science is fun because we get to explore why and how our wires have become crossed, whether that's just because something has been overlooked (e.g. Megaloceros as a powerful, fast runner and not just a pair of giant antlers) or we've been misled by popular culture (e.g. the actual science behind predicting dinosaur vocalisations). I could list articles I've enjoyed researching, writing and illustrating all day but that would be pretty dull for all of us. I'll instead point to the navigation panel on the right that can be used to explore my full catalogue of blog content.
I don't have much new art I can share at the moment, so here's the latest sharable painting I worked on, just so we have some new art content for this anniversary post: it's Yutyrannus huali bellowing on a chilly morning. And yes, it does have a bit of a Christmas card vibe.
It is, admittedly, increasingly difficult to find time to blog as my workload and personal responsibilities have increased in the last ten years. Following the first few years where I was able to post multiple times a month (looking back, I don’t remember ever having that much time on my hands!), I now aim for one post each month, along with regular updates on Facebook, Twitter and Mastodon— do check me out if you’re on the same platforms. I have every intention of keeping the blog going, especially as the line between it and my professional writing is now pretty blurred. Blog posts have become articles and books, and research for books and papers has become blog content. A lot of what you’ve read here has been overspill that I can’t work into other projects, especially from The Palaeoartist’s Handbookan The Art and Science of the Crystal Palace Dinosaurs. At the risk of dipping my hand too far, you’ve also already seen some overflow from my unannounced sixth book — but in which posts, dear reader, which posts?
But I'm saying too much. Sincere thanks to you, my readers, for ten years of blogging fun and especially anyone who’s been around since the beginning. And an even bigger thanks to people who support me at Patreon, without whom I may not be writing here anymore, nor doing any of my other projects, for that matter. Here’s to another decade!
It was ten years ago that palaeoartist John Conway, along with his colleagues Memo Koseman and Darren Naish, published one of the seminal works on palaeoart for the modern age: All Yesterdays: Unique and Speculative Views of Dinosaurs and Other Prehistoric Animals (Conway et al. 2012). Seeing that palaeoartists of the 2000s and early 2010s were already chipping away at the “rigorous reconstruction” conventions established by palaeoartists of the late 20th century, All Yesterdays blew them apart entirely by drawing focus to the “known unknowns'' of restoring fossil organisms. It not only pushed for greater experimentation with style and subject matter, but also, in its cleverest trick, revealed the hilarious/horrific results of applying palaeoartistic techniques to living animals. The slew of online discussions, artworks, memes and projects that followed All Yesterdays have been of variable quality and legitimacy, a comment that applies to the lesser-mentioned crowdsourced follow-up, All Your Yesterdays as anything else, so the legacy of this book is a complex one and there are discussions to be had about its long-term impact. But however we feel about All Yesterdays, we can’t deny that it has shaped much of the conversation around palaeoart in recent years. Its name has, deservedly, become synonymous with the current, postmodern era of extinct animal reconstruction (Witton 2018; Nieuwland 2020; Manucci and Romano 2022).
A decade later, John Conway is back with another book — and this time it’s personal, or, at least, a solo-authored book project, aside from a foreword written by English Literature scholar and dinosaurophile Will Tattersdill. This volume, A History of Painting (with Dinosaurs), imagines what art history may have been like had the great masters of Western art chosen prehistoric animals as their subjects of choice rather than people, landscapes or constructs of human society. It’s hard not to see this, at least partly, as a deep-dive into one of All Yesterdays’ threads about deviating from traditional palaeoart styles. The sense that A History of Painting is a spiritual successor to All Yesterdays also ebbs from its identical size and length, its print-on-demand publishing model, as well as its low price (£19) and sometimes playful, winking tone. But the similarities end there: whereas All Yesterdays used a scattergun approach to critiquing palaeoart in 2012, A History of Painting is a book with a singular point to make.
What that point is, though, is for readers to decide. In John’s own words, “A History of Painting is either a big joke that will make you smile, or a serious questioning of subject matter in art that will make you think”. If A History of Painting is, indeed, a joke, it’s had a long build-up to its punchline. John has been working on this book for two years and produced 50 new paintings specifically for this project, only a few of which can be found online (I've used nearly all of them in this post). They are some of John’s most interesting pieces yet — which is no mean feat, given the quality of his artwork in general— and cement his reputation as one of the most important palaeoartists working today. He has created a series of pastiches of artwork from the 14th to 20th century that range from caricatures of iconic artworks (Mona Heterdontosaurus or The [troodontid] Scream, anyone?) to more “serious” efforts at injecting palaeoart into famous paintings or artistic styles. While a certain amount of Conway DNA exists across each painting, the diversity of styles and genres is seriously impressive and I could easily believe several artists contributed to this book. It’s difficult to think of another palaeoart volume that varies so much stylistically, and I’m including multi-artist compendiums like Mesozoic Art in that consideration. If you’re a fan of Conway palaeoart, you need to grab this for its art alone. I can give no better indication of the quality of artwork than mentioning that I bought a large print of one of the paintings at the launch event (below), and it’s going to be framed and hung in the house somewhere.
Proof, if proof be need be, of my ownership of these Conway Moospods. The animals here are Saltasaurus, rendered over Ploughing in the Nivernais by the 19th century animalier Rosa Bonheur. Art by John Conway, from Conway (2022).
Had A History of Painting only been filled with dinosaur-flavoured riffs on da Vinci, van Gogh or Warhol, I’m not sure I’d be writing about it here. John’s takes on these iconic works are great but we’ve seen so many imitations and parodies of the paintings in question that there’s not much more to be mined from reimagining them, even with dinosaurs. Happily, A History of Painting is mostly comprised of unexpected mashups of paintings and palaeo: a sauropod-themed reimagining of Bacon’s biomorphic Three Studies for Figures at the Base of a Crucifixion, a Late Gothic battle scene retooled with lances swapped out for sauropod necks, a bellowing theropod set against the same lightning flash that frightened Delacroix’s horse. It’s among these that any pretence of the book is a joke falls away, because anything but the most superficial glances at such works gives cause to reflect on dinosaur art more generally. Seeing dinosaurs in such artistic contexts is simply incorrect, but the quality of their superimposition is such that they can’t be dismissed as crude puns at the expense of historic masters. Rather, this juxtaposition of dinosaurs in “serious” artworks gives much to think about palaeoart, our wider attitude to dinosaurs, and maybe even our relationship with nature itself.
Because the book itself is light on text, I found knowing something about the origins of A History of Painting helped my reading of it. Back in 2017, John gave a public talk at a Popularising Palaeontology event about the interaction between dinosaurs and mainstream art, or lack thereof. In a whirlwind tour of the history of painting, John argued that innovations in style have been pursued at the expense of innovations in subject matter, tracking how Western art initially fought to capture the basics of reality, eventually achieving what we’d today describe as hyperrealism or photorealism, and then pushed back to explore new movements like expressionism, surrealism, and abstraction, cumulating in works such as Rauschenberg’s 1951 White Painting. Style, John argued, is now a dead-end for experimentation, with artists having reached beyond reality into the furthest reaches of the abstract. But in terms of subject matter, mainstream artists have remained pretty focused on matters of humanity: our own form and appearance, our religions, beliefs and cultures, our dramas and tragedies, and the local world we inhabit. So if style is dead, maybe the next artistic frontier is… subject matter? And if artists want to continue pushing boundaries and the limits of human experience, what would make better subjects than extinct animals?
The unmistakable painting approach of Gustav Klimt, reworked to feature hadrosaurs. It's recognisably palaeoartistic, but not as we know it. Art by John Conway, from Conway (2022).
It’s this line of thinking that gave rise to A History of Painting. The opening of the book asks, bluntly “did the vast majority of artists really ignore the greatest subject of all, dinosaurs and closely related animals?”. Though approached somewhat facetiously, it's hard not to find some validity here. Consider the wealth of discovery we’ve experienced over the last few centuries: the reality of Deep Time and extinction, the endless parade of exotic organisms, living and fossilised, that exist and have existed on our planet, the vastness of the cosmos and the nature of other worlds, the fundamental components of physical reality… we could go on and on. And yet, these subjects, which represent the very limits of human knowledge and challenge our comprehension of reality and possibility, remain largely untouched by our most famous artists. Those of us that explore the details of the natural world aren’t part of the classic painter canon: we are given different labels (“scientific illustrators”, “wildlife artists”, “palaeoartists” etc.) and our work, if exhibited at all, is more likely to be shown in a natural history museum than the National Gallery.
John is, of course, not the first person to point out the divide between conventional art and palaeoart, nor to imply that palaeoart is undervalued (Mitchell 1998; Lescaze 2017; Manucci and Romano 2020). W. J. T. Mitchell (1998) and Zoe Lescaze (2017) have suggested several reasons for the obscurity of palaeoart, mostly pertaining to stylistic issues and facts of history. Mitchell regarded the work of prominent 20th-century artists as stylistic “throwbacks” compared to trends in mainstream art, while Lescaze argues that the genre draws on too many artistic influences — Romanticism, Impressionism Fauvism and so on — and thus presents a “cacophony of dialects” to scholars attuned to more unified artistic voices and styles. Mitchell further notes that palaeoart developed too late as a genre to capitalise on the animal painting craze of the 19th century.
But there’s a deeper issue, simply identified as “snobbery” by Lescaze (2017). The symbology of dinosaurs is all wrong for refined, dignified artistic traditions. In popular culture, dinosaurs are synonymous with spectacle, violence, mass consumption and childhood interests (Mitchell 1998; Lescaze 2017). Thusly, as Lescze (2017) observes:
“Throw an engraving of an egret above the mantelpiece and no one balks. Hang a painting of a T. rex in the same spot, and the decision screams nerd stuck in second childhood.”
A Monet-esque impressionist take on Tupuxuara. How would you feel about having this hanging over your fireplace? More relevant here, what would your non-pterosaur-fan friends say about it? Art by John Conway, from Conway (2022).
We might thus have some answers to John’s question, the most important being that great artists did not regard dinosaurs as we — as in, the scientists, scholars and enthusiasts who read blogs like this — do today. We think of prehistoric animals as amazing extensions of the living world, species that must have been as majestic and amazing and inspiring as the greatest of today’s creatures. But to non-specialists, these animals are vulgar, monstrous forms represented by tacky merchandise and blockbuster movies. Even if they followed the scientific thinking of the time, artists of the 19th and early 20th centuries would have regarded dinosaurs as inferior animals to mammals and birds (a view that might explain the absence of “lower animals” — invertebrates, reptiles and amphibians — in mainstream painting as well). And, of course, we have the longstanding, entirely false idea that art and science are inherently incompatible. These conspire, Mitchell concludes, to force dinosaurs into a niche well separated from the traditions of studio art:
“...the dinosaur seems to have its ‘proper’ place as the figurehead image of the natural history museum, [where] it helps to reinforce the illusion of a strict separation between nature and culture, science and art. The truth is, this separation is one of cultural status and has absolutely nothing to do with nature, which is just as much the object of art as of science.”
Mitchell (1998), p. 62.
By injecting dinosaurs into classic paintings, A History of Painting continues this discussion in a radical new way, allowing us to explore the inherent “wrongness” of seeing dinosaurs approached as “serious” art subjects. I see John’s book as a direct challenge to the idea that palaeoart must have a purpose, sensu Mitchell’s (1998) comment that “A cubist dinosaur would not be of much use, either to a palaeontologist or to the public” (p. 60). Three cubist paintings (Therizinosaurus, Triceratops and Ankylosaurus) allow us to judge that for ourselves. Are these “useful” paintings of dinosaurs? OK, they distort the appearance of the animals in question, but does a distorted dinosaur serve no purpose? Does bringing a cubist approach to dinosaurs deny us the interpretations we might discuss around conventional cubism, such as its capturing of movement or time, its use of multiple perspectives to convey three-dimensional shapes, and the use of a flat image to capture reality? And, more broadly, it asks why must art of dinosaurs be useful? Can it not be art for the sake of being art, or created purely for aesthetic value? As John pointed out at the A History of Painting launch, the unusual shapes and anatomy of dinosaurs allow for terrific abstractions if we can allow ourselves to abandon the idea that they should only be rendered in ways that show their bodies precisely and clearly. And, ironically, this very discussion already shows that John’s cubist dinosaurs have a purpose, inviting us to question our relationship with dinosaurs in art.
But lest it be thought that A History of Painting is full of strange and bizarre paintings, some works also allow us to recognise the roots of palaeoart itself. John demonstrates that conventional palaeoart has a home, at least stylistically, among the Romantic landscapes of Constable, Corot and Boheur. Here, John creates pieces where dinosaurs are dwarfed by richly painted, detailed surroundings, recalling Henderson-esque landscapes with a Romantic twist. Each could be dropped into a dinosaur textbook without raising eyebrows. His artworks allow for other reflections on palaeoart practises too. His German Renaissance-inspired take on Nemegt Formation dinosaurs allows for a surprisingly effective take on a classic“menagerie” scene with multiple species, the distorted perspective allowing animals of all sizes and shapes to remain visible and uncrowded in what would be an otherwise overly-busy scene. The intentionally warped anatomy of certain restorations provides a new perspective on animal monsterisation: the aforementioned Delacroix Therizinosaurus is disturbing and nightmarish, the reddish hues that accompanied the original facial features of the horse being extended across the face and neck in a fashion that recalls an open wound. And the large number of intimate portraits, often of theropods with forward-facing eyes, encourages us to consider dinosaurs as individuals, a goal we often pursue by adorning them with wear and tear (e.g. scars, scratches, blotches etc.), not quiet, close interaction with viewers. Needless to say, the great stylistic experimentation gives much to ponder about traditional approaches to rendering fossil species. Do some of the paintings in the book transcend “palaeoart” as we might typically define it, or are they "art that features dinosaurs"? Where would we draw the line? Should we even bother with lines at all?
Tarbosaurus hunts Saurolophus in Cretaceous Mongolia, imagined here in Lucas Cranach's 16th Century style. Against all expectations, the strange perspective and lofty point of view make this potentially overcrowded scene very pleasant to look at: it's essentially 5-6 small paintings in one. Art by John Conway, from Conway (2022).
We could go on: there is much to ponder over when thumbing through A History of Painting, and writing this has only prompted even more to think on and discuss. And it’s here, with my brain full of thoughts and ideas, that I find my only real concern (as opposed to criticism) about this project. Much of the above paraphrases points that John has raised in talks about his book, but there’s no substantial discussion of this nature in the book itself. Instead, it leads with a short meta-fiction, suggesting the paintings are recreations of a lost collection of dinosaur artworks by well-renowned artists. The descriptions of the paintings toe this line, making suggestions as to the original artists and subject matter as one might if recreating a series of images from photographs. It’s a fine enough set-up but I worry that it denies the book a context and, dare I say it, an importance that it might otherwise have had, risking it being seen as little more than an exercise in kitsch rather than a considered entry to an ongoing scholarly discussion. A version of A History of Painting was drafted that contained more explanation and text, but didn’t make the cut for being overly stuffy and academic: this is a book designed to appeal beyond a few historians and researchers, after all. I can’t criticise the book for the approach it’s taken because there’s surely no right or wrong way to frame such an unusual project, but I hope its approach doesn’t see it become ignored or brushed off as lightweight frippery. I feel John’s thoughts on this should be recorded somehow, and I wonder if those unused drafts would warrant being turned into a complementary paper or article. And, again, this idea gets my brain turning over: I’m curious to know what non-palaeontological artists and art historians would make of all this. Some sort of public discussion outside of the bubble of palaeo-enthusiasts could be a terrific event. But — no, we must stop.
This wanting the book to be discussed and contemplated is, of course, a strong, if indirect, recommendation for getting yourself a copy. Although it's probably too abstract to have the impact of All Yesterdays, A History of Painting (with Dinosaurs) is a book like no other and another important contribution to the palaeoart book canon from John Conway. Beyond being great to look at, it’s a thought-provoking exploration of not only the value and role of palaeoart, but of our societal relationship with extinct animals and nature, and all for just £19. At the time of writing, there’s still time to get it before Christmas and it should be available pretty much wherever Amazon operates (John has some quick links at his website, but check your national Amazon webpage for details). If you have any stockings to fill for a palaeoart fan, or perhaps for anyone interested in art history, this is well worth your money.
And having mentioned festive season, it’s time at the bar for 2022 at this (now ten-year-old!) blog. However you mark the end of the year, I hope you all have a safe and happy time, and I’ll see you in 2023.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Conway, J. (2022). A history of painting (with dinosaurs). Independently published.
Conway, J., Kosemen, C. M., & Naish, D. (2013). All yesterdays: unique and speculative views of dinosaurs and other prehistoric animals. Irregular books.
Lescaze, Z. (2017). Paleoart: Visions of the prehistoric past. Taschen.
Manucci, F., & Romano, M. (2022). Reviewing the iconography and the central role of ‘paleoart’: four centuries of geo-palaeontological art. Historical Biology, 1-48.
Mitchell, W. T. (1998). The last dinosaur book: the life and times of a cultural icon. University of Chicago Press.
Nieuwland, I. (2020). Paleoart comes into its own. Science, 369(6500), 148-149.
Witton, M. P. (2018). The Palaeoartist's Handbook: Recreating prehistoric animals in art. The Crowood Press.
Mighty Tarbosaurus bataar carries its Therizinosaurus cheloniformis prey through a Masstrichtian forest in the rain. Colour-wise, I've decided that a ruddy-orange dorsum, light venter, disruptive black stripes, rings and large eye masks are meant to hide this 10 m long animal during predatory acts. But without direct palaeocolour data for Tarbosaurus, could we ever evaluate how sensible this colour scheme is?
Today we’re looking at one of the most commonly asked questions about restoring extinct dinosaur appearance: colour. For centuries, queries about the colours and patterns of dinosaurs, and, indeed, most extinct vertebrates, have been effectively non-answerable, save for some arm waving about the merits of camouflage for predation and display patterns for social signalling. Nowadays, advances in analyses and understanding of fossil pigments have allowed us to reconstruct the foundation colours of several dinosaurs in detail, along with those of other popular taxa like pterosaurs and marine reptiles (see Vinther 2015 and Smithwick and Vinther 2020 for overviews). This new frontier in dinosaur science has helped to flesh out not only the life appearance of dinosaurs, but also their ecology: their habitat preferences, their daily activity patterns, their predation concern and so on (e.g. Vinther et al. 2016).
Deducing dinosaur colour to this level of precision requires exceptionally high-quality preservation of their skin, down to the microscopic level, so that their pigment cells (melanosomes) can be identified. Unfortunately, this excludes the vast majority of dinosaur specimens from such analyses. Dinosaur skin is not only rare, but often occurs as mere sediment impressions rather than films of geochemically-preserved organic matter. This preservation style applies to a great number of the most famous dinosaurs so, unless some radical new science finds a way to assess colour from skin texture alone, the colours of our favourite extinct saurian taxa will probably be lost to time forever.
But can we tackle this problem from another angle? In recent decades, biologists have made enormous strides in understanding living animal colouration, looking at how it relates to habitat preferences, camouflage, signalling behaviour, body size, posture, visuality acuity and so on. Has the science around modern animal colouration advanced to the point where we can start to make tighter predictions about the colours of extinct animals? I regard this as an important question because, as much as our depictions of dinosaur anatomy have tightened since the late 20th century, our application of colour is still pretty lawless, even among professional palaeoartists. We present the same animals with colour schemes that are totally adaptively opposed to one another — one artist’s vivid blue hadrosaur is met with another’s muted browns and reds — and yet they’re both meant to be of equal scientific credibility. But how can that be so? Colours and patterns are generally thought to be under the same adaptive pressures as other parts of animal anatomy and thus should correlate, to a greater or less extent, with aspects of behaviour and ecology. There probably is, at some level, a "right" and "wrong", or at least a "likely" and "less likely" aspect to colour restoration, just as there is with all other aspects of palaeoartistry. But how can we evaluate this without palaeocolour data? Enter, stage left, the last two decades of studies of living animal colour. Can they help constrain, even in a general way, our efforts at colouring animals from Deep Time?
What flavour Australovenator wintonensis is your favourite — red, green, blue or orange? It's strange that, as consistent as we're getting with depicting dinosaur anatomy, you could present any one of these contrasting colour variants with equal scientific validity, even though they each imply very different interpretations of Australovenator biology.
Time for a case study
To investigate this, I thought we could look at a well-known group of dinosaurs to see what, if anything, living animals might suggest about their colouration. As you’ve guessed from the article title, we're using big predatory theropods for this exercise, for several reasons: 1) they’re a popular art subject, so this article should be of wide interest; 2) as regular readers will have worked out, I’m currently involved in a few big theropod projects so have been drawing them fairly continuously for a while now; and 3), the biology and ecology of big theropods are comparatively well-researched, and that helps when plugging fossil data into models of extant animal colour. And yes, we could restrict this to a more specific theropod clade but, from what I know about giant predatory dinosaurs, I’m not sure the conclusions we’d draw for big allosauroids would be much different to those of tyrannosaurines or large megalosauroids. If we’re sticking to what we know about these animals, not what we speculate and imagine about them, they only offer so much data to compare against living species.
There are plenty of caveats with this comparison, of course. No living creature is ecologically or phylogenetically close to the largest Mesozoic theropods, and our modern environments are different to those inhabited by our case study subjects. But we might also consider the importance of uniformitarianism, the adage that “the present is the key to the past”. We can’t say whether modern animals are perfect models for the colour of Mesozoic species, but they offer the only large, statistically-viable sample size of biological colour for us to work with. We are surely better off making informed guesses about extinct animal appearance using modern species as a guide, dodging known pitfalls where we can, than simply speculating wildly.
More worrying than concerns about comparing the past with today is that the controlling factors of animal colouration are extremely complicated, and it’s not clear how we can account for this. Indeed, for all of our science and ideas around animal colour, we still have lots to learn about it. Many popular, widely communicated interpretations of animal colours and patterns are only now being experimentally evaluated (Caro 2005), which means we are still struggling to understand some foundational aspects of certain colour schemes (Caro 2013). This is especially the case for predatory species, the colours of which have been relatively unexplored compared to those of prey animals (Pembury Smith and Ruxton 2020). To that end, we must temper our expectations. As neat as it would be to pour details like extinct animal size, habitat preference and trophic level into an algorithm to receive — *ping!* — a series of likely colours and patterns, our conclusions here, if any, are going to be of a more generalistic, broader nature.
Camouflage and detectability in large living predators: what does it mean for big theropods?
For all the new work that’s been done on animal colour, we still recognise that the principal pressures on animal colouration are essentially what Darwin observed in his 1871 book The Descent of Man. This is a conflict between natural selection, which promotes colour configurations that help animals remain undetected by predators, avoid temperature stress and generally survive from day to day, and sexual selection, which promotes the adoption of bold, broadcasting colours and patterns that attract mates and deter social rivals. So the first thing we might explore for big Mesozoic dinosaur predators is how our largest living terrestrial carnivores express this conflict: are they more concerned with basic natural functionality or sexual signalling? We're specifically interested in our giant theropod ecological analogues here: big animals that hunt and kill relatively large prey items. Predators that subsist on smaller, bite-sized animals don't qualify, because their ecology isn't sufficiently similar.
A selection of the largest predatory animals of modern times and their camouflage schemes, universally showing a strong adaptive emphasis on concealment regardless of habitat type, phylogeny or locomotor method. A, Ora, or Komodo dragon Varanus komodoensis (background matching); B, lion, Panthera leo (background matching); C, tiger Panthera tigris (disruptive colouration); D, polar bear Ursus maritimus (background matching); E, saltwater crocodile Crocodylus porosus (background matching); F, golden eagle Aquila chrysaetos (background matching); G, great white shark Carcharodon carcharias (countershading). All images from Wikimedia: A, Yuliseperi2020, CC BY-SA 4.0; B, Bernard DUPONT from FRANCE, CC BY-SA 2.0; C, Charles J. Sharp, CC BY-SA 4.0; D, Andreas Weith, CC BY-SA 4.0; E, fvanrenterghem, CC BY-SA 2.0; G, Juan Lacruz, CC BY-SA 3.0.
Across vertebrate groups, and across habitat types, our biggest modern predators are pretty consistently (maybe entirely consistently) primarily coloured for concealment: that is, they have camouflaging colours and patterns which hide their presence from their prey. This applies as much to mammals, which are a relatively drab group overall on account of several ecological and physiological factors (Caro 2013), as it does to clades that have the adaptive capacity to produce the most brilliant and striking colour schemes in nature, such as lizards, snakes, birds and fish. So maybe that’s our first note: big predators in the modern day are all about cryptic colouration, with little in the way of conspicuous display patterning.
Research on the impact of body size on predatory ecologies sheds light on why big predators seem to be consistently camouflage-coloured, and it’s a simple explanation: bigger animals are generally more conspicuous than smaller ones, even when they're trying their best not to be seen. The relationship between predator size and concealment capacity is still being investigated but a trend between size and conspicuousness seems to apply widely across Animalia, even in species with famously adept camouflage adaptations, like chameleons (Cuadrado et al. 2001; Pembury Smith and Ruxton 2020). Size doesn’t just affect detectability, either: it also correlates with prey response. Bigger predators instigate more vigorous reactions than smaller ones, such that prey species react sooner, flee further, or initiate more aggressive counter-responses (Stankowich and Blumstein 2005). There are strong pressures, therefore, on big predators to do what they can to remain hidden. Their size already puts them at a disadvantage for stealthily approaching prey, and they are going to have to run further or fight harder once they give up their hiding spot. Given that the largest theropods are the biggest terrestrial predators that have ever lived, we have to wonder what this link between body size and cryptic capacity implies for their colouration. Is one obvious inference that big theropods needed all the help they could get to remain inconspicuous? Would predators already handicapped by their greater detectability and exaggerated prey responses really have some of the signalling-dominant, hyper-obvious colour schemes we've given them from time to time?
Giganotosaurus adapted for the open county with high-bodied, sharply marked countershading, from my recent post about the possible facial anatomy of this animal. But note the ornament on this animal's head: I feel I gave it a pretty meaty set of soft-tissues around its snout and eye, but Giganotosaurus is still pretty undecorated compared to some theropods. Is this something we can read into — does the extent of cranial ornament tell us something facial colouration?
While fossils do not tell us anything about this correlation directly, I wonder if some anatomical evidence points to larger predatory dinosaurs aiming to be less conspicuous. Mid-and large-sized theropod fossils tend to have bony cranial ornaments more often than smaller ones (Gates et al. 2016), but in my estimation (by which I mean, this hasn't been verified by any study), the ornamentation in very large species is generally reduced and less spectacular than that of their smaller cousins. In tyrannosauroids, for instance, we see a general shift away from tall midline cranial crests in smaller, earlier species towards low-relief rugose surfaces, small horns or blunt bosses in larger taxa (Gates et al. 2016). Indeed, the very largest theropods are some of the dullest-looking, at least in terms of cranial ornament. Consider the flattened orbital bosses and rostral rugosities of Tyrannosaurus and Tarbosaurus, or the low, corrugated textures over the snouts of giant carcharodontosaurids. We can only speculate on what impact these ornaments might have had on theropod camouflaging efforts, but it’s well-established that distinctive body outlines can increase detectability, to the extent that modern predators attempt to hide them from their prey where possible (see below).
Whatever their adaptive significance, these reduced facial ornaments give us grounds to think about cranial colouring. Faces are often sites for signalling patterns and colours in modern species (e.g. Caro et al. 2017) and a reduction in bony facial ornament could indicate a lessened emphasis on this behaviour, possibly including muted facial colouration. A caveat here is that elaborate osteological features are only ever suggestive of striking colours and patterns, not directly correlated. But part of the palaeoart game is looking for clues about the nature of these animals wherever we can, and an absence or reduction of showy features is something we can factor into the reasoned speculation we must utilise when creating colour schemes.
I don't think we make enough of how display-adapted Spinosaurus aegyptiacus was, and how weird that is for not only a giant dinosaur predator, but any giant predator. Here, a gaggle of Spinosaurus show off their sails and tails, display structures (well, probable display structures, in the case of the tail) that use almost every inch of their axial length for showing off. So how does this fit into your "big theropods were camouflage-colour dominant" narrative, smart guy?
The elephant in the room here, of course, is Spinosaurus, which is highly unusual for being a giant apex predator with the same tailor as a peacock. This was a carnivore with an unprecedented disregard for remaining inconspicuous or having an anonymous body profile. For all the controversy over this animal, one aspect we all agree on is that its enormous sail was a sociosexual display device (see Hone and Holtz 2021 for references and discussion). Doesn’t this doesn’t torpedo the wider point being made here about predator size and possible camouflage needs? On the contrary, it might support it. As something straddling the terrestrial-aquatic realm, normal rules about camouflage and crypsis may not have applied to Spinosaurus. We see this evidenced in modern times in that the "rules" of camouflage in terrestrial settings are not the same as those of aquatic habitats (Caro 2013), and we should probably allow for, or even expect, some weirdness from animals operating at that interface. The atypical ecology of spinosaurids may have liberated them from the adaptive pressures experienced by purely terrestrial dinosaur predators, allowing them to become more ornamental and spectacular. Perhaps the fishy prey of Spinosaurus barely saw the full outline of their largest predator, an especially viable idea in the (I think, superior) “giant heron” ecological model favoured by several authors (e.g. Hone and Holtz 2021; Sereno et al. 2022).
Pigment availability
Moving on, can we get a sense of the skin pigmentation available to giant theropods, thus letting us know which paints/colouring pencils/digital palettes to crack open? Here, we have to think about the availability of environmental pigments, like carotenoids. Many readers will know that animals cannot create all the pigments used in their integument and that some are obtained through eating plants or microbes. Carotenoids and other environmental pigments create some of the most vivid colours seen on animals today, including hot reds, bright oranges and canary yellows. But environmental pigments are hard to source in terrestrial settings, to the extent that even tiny songbirds compete with one another to source them (Blount 2004; Biard et al. 2005). Outside of specialist ecologies, the most famous being that of flamingoes, larger terrestrial animals tend to make do with pigments they can manufacture themselves, such as melanin. This is one reason why so many terrestrial animals are earthy tones, such as greys, blacks, browns, orange-reds, and white (where pigment is withheld). But structural colour, features of skin, scales and feathers that manipulate light to create colour without pigmentation, has also been developed across all vertebrates and is exploited to produce greens and blues. In all probability, it’s from these basic pigment and structural palettes that giant theropods were deriving their hues. Unless conditions of the past were very different to those of today, it’s hard to imagine multi-tonne terrestrial animals finding enough carotenoids to develop large patches of particularly intense pigmentation.
Specifics of patterning
Our discussion raises a notch in complexity as we move to consider giant theropod skin patterns, even if we stick within the camouflage-dominant framework outlined above. Concealment strategies are adapted to specific habitats, predation styles and prey types because no one system is universally effective. Indeed, one of the few constant rules of camouflage — that, no matter how perfectly a crypsis strategy works on a stationary animal, movement always gives the game away (Pembury Smith and Ruxton 2020) — is of little use to us here because we don’t know where and how big theropods hunted. The concealment strategy of an endurance predator, one that simply hounds its prey tirelessly, waiting for it to become vulnerable from exhaustion, might be different to that of an ambush predator that relies on surprise, springing at its prey at the last moment for a short chase.
These are only the first factors to consider. Predator colours are also modified by the time of day the predator tends to operate, as well as their position in the food chain: some have to be worried about being prey items themselves. And that, in turn, is altered by the colour schemes that can be created by different integument types (e.g. fibres vs. naked skin vs. scales), as well as the functional impacts of pigmentation. Darker pigments, for instance, can protect skin from harmful UV rays and may have antibacterial properties but, conversely, also absorb more solar heat and increase an animal’s thermal load (Walsberg 1983; Caro 2005; Caro and Mallarino 2020). There’s a lot to think about here, and the fact we still can’t account for these and other variables reliably in living animals is why biologists still consider our knowledge of animal colour to be fairly limited. It goes without saying that, if we’re still working out what’s happening among living species, robust predictions of camouflage patterning in extinct animals are way off.
The colour schemes we give our dinosaurs have functional implications beyond interacting with other animals. This dark, adult Tyrannosaurus would be well-protected from solar radiation by its dark skin, but it would absorb a lot of heat in direct sunlight.
Nevertheless, we may be able to narrow down some possibilities for giant theropods by looking at what works for large modern predators. Most employ background matching, where their skin tone approximates that of their surroundings, or else they use countershading, where dark upper regions and lighter undersides disrupt the formation of shadows, diminishing contrast with the background (note that this is disputed by some, there is actually a fair amount of controversy around countershading function: see Ruxton et al. 2004; Rowland 2009). Other predators use disruptive colouration, where high-contrast colours break up body outlines and disguise distinctive features such as eyes. Unlikely strategies for big theropods are masquerading tactics: attempts to match unexciting objects like rocks or twigs. To pull off this illusion, masqueraders have to resemble something of equivalent size and shape, and that becomes harder at larger sizes, perhaps explaining the absence of this method among large terrestrial predators today. This strategy is distinct from mimicry, where an organism adopts the appearance of another species to be misleading about its true nature (Skelhorn et al. 2010).
With several patterning options on the table, progressing further with this discussion is only possible if we start making assumptions about giant theropod ecology, pushing us further into the realm of inference and speculation. But we can ground ourselves by considering the results of studies into camouflage function and performance. For instance, if countershading does indeed work to disrupt shadowing, then studies show that a sharp, high-body colour transition would work better in an open setting than a more gradual colour change lower on the flank, which obscures animals more effectively in forested settings (Vinter et al. 2016). We generally see more uniform, low-contrast colours on big animals in open habitats because large patches of colour generally don’t conceal animals as well in woodlands (Pembury Smith and Ruxton 2020, although flat-grey elephants are reportedly remarkably difficult to find once they enter forests — see Caro 2013). Conversely, high-contrast patterns seem to work better at hiding animals in vegetated or otherwise busier environments.
Baby tyrannosaurs, barely a metre long, with cryptic colours that help them blend into the forest floor. In all likelihood, baby tyrants were at high predation risk and it seems reasonable to assume they used camouflage tactics to avoid being eaten. But the colour schemes of infants may not have worked so well for their gigantic parents, nor even older juveniles or subadults. Might tyrannosaurs and other giant theropods have tracked through multiple colour morphs en route to somatic maturity?
We can consider things like the age of our restoration subjects, too. In scaly animals (the only skin type we currently have direct evidence for in giant predatory theropods, even if we can’t rule out the possibility of some protofeathering), colour vividness tends to reduce with age (Olsson et al. 2013). This change may not just be physiological, but also adaptive. The juveniles of all animals, including apex predators, are targetted by carnivores and their colouration has to be multi-functional, hiding them from predators as well as — in precocial species — their prey. This is often achieved with disruptive patterning. Stripes, spots, bars and other features may serve an additional role, achieving a “motion dazzle” effect that confuses predators about animal speed and direction, or draws focus to less critical anatomies, like tail tips (Murali & Kodandaramaiah 2016). Dazzling capacity diminishes at lower speeds and agility, and is thus less useful in larger animals (Pembury Smith and Roxton 2020), perhaps partly contributing to the dulling of living reptiles as they approach adulthood. We should not imagine that juvenile theropods transitioned to their adult colours straight away, however. It took decades to grow gigantic theropodan predators and, in all probability, the route to adulthood was via several different ecological niches (e.g. Holtz 2021), each of which may have had different adaptive pressures on colouration. So maybe giant theropods had several colour schemes throughout their lives, and we should render them as being colour-adapted to their various age-specific lifestyles? We could go on listing the adaptive aspects of different animal skin patterns all day, but you get the idea. There's a lot of camouflage science we could factor into our reconstructions, even if we can't ever know the real colours and patterns of our subject species.
So... does animal colour science help us in palaeoart?
Background-matching, age-dulled Tyrannosaurus rex takes on a countershaded, partly disruptively-coloured Edmontosaurus annectens. These guys are mainly here to stomp about and wake people up with some Hardcore Dino Action™ in case anyone has drifted off when reading this long, long post.
Let’s conclude by returning to our main question: can studies of living animals constrain our speculations about the colours of dinosaurs, or will colour restorations forever remain a crapshoot when we don’t have palaeocolour data? Here, we've extrapolated the findings of predator-specific colour studies to giant, terrestrially-hunting theropods and, based on these, we've suggested that large dinosaur predators...
were likely under very strong pressures for crypsis
probably didn't load their skin with many environmental pigments
likely expressed background matching, countershading or disruptive patterning, depending on their specific ecologies
may have had several colour schemes throughout their lives as their ecology changed with age
Down the line, we can discuss the merits of these predictions — do remember that you're reading a secondary take on all the science discussed here and that you may come to different interpretations based on your own literature crawl. But it's not these specific findings that are most important here. Rather, it's that this case study shows what animal colour science can offer to the process of restoring one type of extinct animal, as well as its broader potential for focusing our loosely-constrained applications of colour within palaeoart. The points made above, or others like them, do not give a colour scheme for giant theropods, but they do suggest that some concepts are more likely than others, and even rough guidance isn't to be sniffed at when we're otherwise running virtually blind. It's strange that palaeoartists are often able to point to core palaeontological studies for interpreting fossils, and core anatomical studies for depicting anatomy, but we don't generally talk about or know the same literature on animal colour. I wonder if it'll eventually be worth keeping up with developments in this field as much as we do new fossil and anatomical data — if we’re not at this point already. The result can only be more scientifically credible and realistic artwork, and that's a win for everyone.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Biard, C., Surai, P. F., & Møller, A. P. (2005). Effects of carotenoid availability during laying on reproduction in the blue tit. Oecologia, 144, 32-44.
Blount, J. D. (2004). Carotenoids and life-history evolution in animals. Archives of biochemistry and biophysics, 430(1), 10-15.
Caro, T. I. M. (2005). The adaptive significance of coloration in mammals. BioScience, 55(2), 125-136.
Caro, T. I. M. (2013). The colours of extant mammals. In Seminars in Cell & Developmental Biology (Vol. 24, No. 6-7, pp. 542-552). Academic Press.
Caro, T., & Mallarino, R. (2020). Coloration in mammals. Trends in Ecology & Evolution, 35(4), 357-366.
Caro, T., Walker, H., Santana, S. E., & Stankowich, T. (2017). The evolution of anterior coloration in carnivorans. Behavioral Ecology and Sociobiology, 71, 1-8.
Cuadrado, M., Martín, J., & López, P. (2001). Camouflage and escape decisions in the common chameleon Chamaeleo chamaeleon. Biological Journal of the Linnean Society, 72(4), 547-554.
Gates, T. A., Organ, C., & Zanno, L. E. (2016). Bony cranial ornamentation linked to rapid evolution of gigantic theropod dinosaurs. Nature Communications, 7(1), 12931.
Holtz Jr, T. R. (2021). Theropod guild structure and the tyrannosaurid niche assimilation hypothesis: implications for predatory dinosaur macroecology and ontogeny in later Late Cretaceous Asiamerica1. Canadian Journal of Earth Sciences, 58(9), 778-795.
Hone, D. W., & Holtz Jr, T. R. (2021). Evaluating the ecology of Spinosaurus: Shoreline generalist or aquatic pursuit specialist?. Palaeontologia Electronica, 24(1), 1-28.
Murali, G., & Kodandaramaiah, U. (2016). Deceived by stripes: conspicuous patterning on vital anterior body parts can redirect predatory strikes to expendable posterior organs. Royal Society Open Science, 3(6), 160057.
Olsson, M., Stuart-Fox, D., & Ballen, C. (2013). Genetics and evolution of colour patterns in reptiles. In Seminars in cell & developmental biology (Vol. 24, No. 6-7, pp. 529-541). Academic Press.
Pembury Smith, M. Q., & Ruxton, G. D. (2020). Camouflage in predators. Biological Reviews, 95(5), 1325-1340.
Rowland, H. M. (2009). From Abbott Thayer to the present day: what have we learned about the function of countershading?. Philosophical Transactions of the Royal Society B: Biological Sciences, 364(1516), 519-527.
Ruxton, G. D., Speed, M. P., & Kelly, D. J. (2004). What, if anything, is the adaptive function of countershading?. Animal Behaviour, 68(3), 445-451.
Sereno, P. C., Myhrvold, N., Henderson, D. M., Fish, F. E., Vidal, D., Baumgart, S. L., ... & Conroy, L. L. (2022). Spinosaurus is not an aquatic dinosaur. Elife, 11, e80092.Skelhorn, J., Rowland, H. M., Speed, M. P., & Ruxton, G. D. (2010). Masquerade: camouflage without crypsis. Science, 327(5961), 51-51.
Smithwick, F., & Vinther, J. (2020). Palaeocolour: A History and State of the Art. In: Foth, C., Rauhut, O. (eds) The Evolution of Feathers. Fascinating Life Sciences. Springer. 185-211 pp.
Stankowich, T., & Blumstein, D. T. (2005). Fear in animals: a meta-analysis and review of risk assessment. Proceedings of the Royal Society B: Biological Sciences, 272(1581), 2627-2634.
Vinther, J. (2015). A guide to the field of palaeo colour: Melanin and other pigments can fossilise: Reconstructing colour patterns from ancient organisms can give new insights to ecology and behaviour. BioEssays, 37(6), 643-656.
Vinther, J., Nicholls, R., Lautenschlager, S., Pittman, M., Kaye, T. G., Rayfield, E., ... & Cuthill, I. C. (2016). 3D camouflage in an ornithischian dinosaur. Current Biology, 26(18), 2456-2462.
Walsberg, G. E. (1983). Coat color and solar heat gain in animals. BioScience, 33(2), 88-91.
The hero of the hour, Triceratops horridus. But how often were those long horns stuck into predatory dinosaurs in defensive action? I feel a long discussion coming on...
A persistent idea around dinosaur biology is that the ceratopsids, or horned dinosaurs, were among the most formidable prey for predatory theropods. Though lacking armour, the large cranial horns and frills of these dinosaurs have been widely interpreted as having anti-predator potential, functioning like a knight’s lance and shield in their capacity to stab and parry attacking carnivores. Such notions are well over a century old and have taken on a life of their own in palaeontological media, especially thanks to widespread romanticising of the relationship between Triceratops and Tyrannosaurus, genera that we’ve decided represent the ultimate expression of dinosaurian predator and prey. Henry Fairfield Osborn wrote on such matters as early as 1917:
“The first of these [dinosaurs with anti-predator anatomy] are the aggressively and defensively horned Ceratopsia, in which two or three front horns evolved step by step, with a great bony frill protecting the neck. This evolution took place stage by stage with the evolution of the predatory mechanism of the carnivorous dinosaurs, so that the climax of ceratopsian defense (Triceratops) was reached simultaneously with the climax of Tyrannosaurus offense. This is an example of the counteracting evolution of offensive and defensive adaptations, analogous to that which we observe today in the evolution of the lions, tigers, and leopards, which counteracts with that of the horned cattle and antelopes of Africa, and again in the evolution of the wolves simultaneously with the horned bison and deer in the northern hemisphere.”
Osborn 1917, p. 224-225.
At this early stage in dinosaur research, the likes of Triceratops and Tyrannosaurus weren't viewed as laudable champions of evolution, but as animals so stupid and instinct-driven that their predatory and anti-predatory strategies had to be as simple and idiot-proof as possible. Tempting as it is to reduce this passage from William Matthew’s 1915 American Museum of Natural History book Dinosaurs to some choice soundbites, it’s such an amazing window into old-school concepts of dinosaurian stupidity that I present it here in its full glory.
“[Tyrannosaurus] probably reached the maximum of size and of development of teeth and claws of which its type of animal mechanism was capable. Its bulk precluded quickness and agility. It must have been designed to attack and prey upon the ponderous and slow moving Horned and Armored Dinosaurs with which its remains are found, and whose massive cuirass and weapons of defense are well matched with its teeth and claws. The momentum of its huge body involved a seemingly slow and lumbering action, an inertia of its movements, difficult to start and difficult to shift or to stop. Such movements are widely different from the agile swiftness which we naturally associate with a beast of prey. But an animal which exceeds an average elephant in bulk, no matter what its habits, is compelled by the laws of mechanics to the ponderous movements appropriate to its gigantic size. These movements, directed and controlled by a reptilian brain, must needs be largely automatic and instinctive. We cannot doubt indeed that the Carnivorous Dinosaurs developed, along with their elaborately perfected mechanism for attack, an equally elaborate series of instincts guiding their action to effective purpose; and a complex series of automatic responses to the stimulus afforded by the sight and action of their prey might very well mimic intelligent pursuit and attack, always with certain limits set by the inflexible character of such automatic adjustments. But no animal as large as Tyrannosaurus could leap or spring upon another, and its slow stride quickening into a swift resistless rush, might well end in unavoidable impalement upon the great horns of Triceratops, futile weapons against a small and active enemy, but designed no doubt to meet just such attacks as these. A true picture of these combats of titans of the ancient world we cannot draw; perhaps we will never be able to reconstruct it. But the above considerations may serve to show how widely it would differ from the pictures based upon any modern analogies.”
The image that launched a thousand Cretaceous daydreams... but not the version you know. This is the rarely-seen, 96 cm wide (presumably preparatory) version of Charles Knight's classic 1928 Triceratops vs. Tyrannosaurus mural, held today at Princeton University Art Museum. Knight's near-blindness meant that he could only execute the mural at scale; other artists then painted the better-known, full-size version. I don't need to explain why this image is included in this post.
Over the last century, our views on dinosaur physiology and intelligence have (perhaps thankfully) changed, but the concept of horned dinosaurs protecting themselves with their facial ornaments has not. Some authors (e.g. Colbert 1948) have seen anti-predator functions as the primary role of ceratopsid horns, and even those who view these structures as evolving under different selection regimes (e.g. Hone et al. 2011) assume some predator defence was possible. Seminal figures like Robert Bakker have worked sweeping hypotheses from concepts of long-standing predator-prey interactions between dinosaur species, rephrasing Osborn’s “counteracting evolution of offensive and defensive adaptations” into the catchier “Mesozoic arms race” (Bakker 1986). Of course, Triceratops and Tyrannosaurus are considered the final, ultimate example of this era-spanning feud. On their relationship, Bakker wrote:
“No matchup between predator and prey has ever been more dramatic. It’s somehow fitting that these two massive antagonists lived out their co-evolutionary belligerence through the very last days of the very last epoch in the Age of Dinosaurs.”
Bakker 1986, p. 240.
Gregory S. Paul, also a fan of the idea that Triceratops was the apex challenger to theropod aggressors (Paul 1988), has further worked the concept of ceratopsid predatory combat into other hypotheses. Specifically, in 2008 he proposed that such dangerous prey items were a factor in the short (c. 30 year) lifespans of tyrannosaurids, while also echoing Bakker’s concept of an “arms race” between these clades (“the upgrading the weaponry in tyrannosaurids and ceratopsids… may represent a Red Queen arms race”; Paul 2008, p. 344.).
Today, a large body of evidence has challenged the idea that ceratopsid evolution was driven by developments in predatory dinosaurs. Instead, it points to ceratopsid skulls being primarily shaped by their own intraspecific behaviour. First proposed in the 1970s, this concept arose after researchers noted the many similarities between the sexually-selected horns of living animals and the ornaments of horned dinosaurs. These shared features include their exaggerated and often complex shapes, their functional peculiarity (i.e. that many seem maladapted for other activities, including predator defence), their positive allometry (that they grow faster than the rest of the skull), their high amount of intraspecific variation, and their high morphological diversity between species (e.g. Farlow and Dodson 1975; Spassov 1979; Sampson et al. 1997; Horner and Goodwin 2006, 2008; Hone et al. 2011, 2016; Knell et al. 2012). Evidence that horned dinosaurs injured each other in ways consistent with modelled ritualised combat (e.g. Farke 2004; Farke et al. 2009; D’Anastasio et al. 2022) supports the hypothesis that their horns were employed against one another, not necessarily other dinosaurs, and, along with their fossilisation in huge monospecific bonebeds, we can readily rationalise horned dinosaurs as boisterous animals with somewhat bovid-like behaviours.
Centrosaurus aperatus, a ceratopsid that's so familiar nowadays as to seem unremarkable. But look at that face anew, dear reader: what a crazy animal. Skulls like those of ceratopsids are ripe contenders for dinosaur anatomy shaped by sexual, or at least intraspecific, selection pressures.
The concept of ceratopsid horns serving primarily as anti-predator devices idea has not, however, entirely been set aside despite these data. Nowhere is this more obvious than in popular culture, where horned dinosaurs frequently employ their ornament in life-or-death struggles against predators, and we routinely discuss “armed” dinosaurs as being more dangerous prey than their "unarmed" relatives. But are these action-packed scenarios really a defensible alternative to their horns being used in intraspecific display and aggression? How realistic, really, are these predatory scenarios? Rather than taking the tried and tested route to address this by looking into ceratopsid skull form and function, we’re going to look at the use and evolution of horn-like structures (horns, antlers, ossicones etc.) in living vertebrates. Our view on extinct animal behaviour, after all, is seen through the lens offered by living species and it’s generally our assessment of modern taxa that dictates behavioural models for extinct animals, not the other way around. The anti-predator behaviour of living animals, even just those with horns, is a huge topic that is far too broad and multi-faceted to cover in detail here — especially as I need this to be a relatively short article* — but even in this brief visit, I hope we can hit a few key points that may give food for thought on horn function in Mesozoic animals.
*Yeah, nice try, Past-Mark.
An obvious place to begin is with well-known examples of horn-like structures being used as predator deterrents. It’s absolutely true that some species, like muskox, African buffalo, various rhinos and red deer use their cranial ornament aggressively against predators (Geist 1966, 1999; Schaller 1972; Kruuk 1972) and it is assumed that predator deterrence may explain the presence of horns in a great number of bovids (e.g. Packer 1983; Bro-Jørgensen 2007; Stankowich and Caro 2009; Metz et al. 2018). However, the idea of widespread horn use against predators has been challenged because field observations show such behaviour is rare among many species, and often of limited effectiveness (Estes 1991; Roberts 1996; Gerstenhaber and Knapp 2022). Some groups, like antelopes, are rarely or never witnessed using their sometimes enormous horns in defence against attacking carnivores, even when faced with certain death (Schaller 1972). Most deer seem to behave in a similar fashion, preferring to run or hide from predators despite their capacity to gore and kill conspecifics with their antlers. Indeed, there are indications that antlers may have a deleterious effect on prey species, with Geist (1966) reporting that antlerless moose are more capable opponents against wolves than their "armed" relatives. This may not be the case for all deer, however, with American elk proving more vulnerable to predators once their antlers are cast (Metz et al. 2018). But sometimes losing cranial weaponry makes no difference to predator vulnerability at all, as is the case for black rhinoceros. The necessary act of dehorning these animals to deter poachers shows that both adult and calf survivability are little affected by the removal of their horns (Chimes et al. 2022), suggesting that these structures have a non-essential role in thwarting predatory efforts.
When discussing anti-predation strategies involving horns, the African buffalo Syncerus caffer is one of the go-to species. And yet, these large, formidably armed animals are some of the preferred prey of lions, and are sometimes subdued by single individuals. Photo from Wikimedia, by Diamond Glacier Adventures, CC-BY 2.0.
Perhaps against expectation, not all animals with horn-like structures employ them in defence. Moose generally kick attackers, a behaviour they share with giraffes, who also lash out at predators with their powerful legs rather than bludgeoning them with their armoured, tri-horned heads (Gesit 1966, 1999). Kicking strategies are, of course, also available to species that we might mistake for being “defenceless” from their lack of horn-like anatomy. In some cases, these animals can be far more aggressive than their better-armed contemporaries. Horses, especially zebras, exhibit pronounced anti-predator aggression where they bite and kick attacking cats and hyenas (Kruuk 1972), and with such force that they may explain sightings of lions with shattered jaws (Schaller 1972). Zebras are also recorded as charging towards predators in a fashion that neutralises predatory effort. I like ecologist George B. Schaller’s account of this behaviour where he describes lions simply watching zebras running at them, as any effort to grab them would be “like jumping on a fast-moving train from a standstill” (Schaller 1972, p. 265).
We can augment our discussion further by switching our focus from prey species to their predators. If cranial weapons are effective predator deterrents, we might expect predators to avoid species with horn-like structures or, at least, those more likely to wield them aggressively. And yet, prey preferences seem largely determined by the energy investment demanded in animal capture (Schaller 1972) rather than the presence or absence of cranial armaments. Pouring cold water on romantic notions of life-and-death battles of horns and hooves vs. claws and teeth, field ecology suggests that large predators preferentially target species that are abundant, live in dense populations, and are of a size that provides a suitable reward against the effort of capture. Potential prey species are more likely to be ignored because they are too small, and thus do not provide enough nutrients for the predatory effort, or are too big, and will thus require an unreasonable degree of energetic investment to bring down. This is not to imply that the threat of injury isn’t factored into these behavioural calculations, but we just don’t routinely see predators avoiding horned, aggressive prey in modern ecosystems. On the contrary, both spotted hyenas and lions routinely attack African buffalo (lions especially), a species which is very well known for its horn-led predator defence (Kruuk 1972; Schaller 1972).
Clearly, the idea that horn-like structures serve as anti-predator devices is complicated by a lot of conflicting data. While no one doubts that these anatomies are sometimes used to deter predators, zoologists are engaged in a long-running scientific conversation about the extent and significance of their anti-predator role (e.g. Geist 1966; Estes 1991; Roberts 1996; Caro et al. 2003; Gerstenhaber and Knapp 2022). One especially important issue, which has implications for our discussion of dinosaurs, is whether the horn-like structures of female mammals exist primarily to deter predators (Bro-Jørgensen 2007; Stankowich and Caro 2009). While the formidable cranial weaponry of male mammals is often readily explainable through sexual selection (on which, see below), the function of the same anatomies in females is harder to fathom. Some argue that predator defence is their main purpose, which would imply a much wider role for this behaviour than has been documented in field studies. But others point to social selection as their principal adaptive driver (e.g. to dispute territories or mimic males) or simply regard them as non-functional, suggesting they only exist at all because of a genetic link to male horns. The latter must be the case for the females of some species, such as giraffes, which reportedly never seem to do much of anything aggressive with their heads.
I really like this image from Nikolay Spassov's (1979) paper on horned dinosaur evolution for its novel depiction of horned dinosaur combat. I especially like the interlocking of the frill spikes as we (or, at least, I) tend to forget about them possibly playing a role in physical competition. The horn-like structures of male mammals are shaped to match certain styles of intraspecific combat, and it's possible that ceratopsids were driven by similar evolutionary forces.
Thankfully, we can push most of these conflicting ideas and caveats aside to discuss the horn-like structures of male mammals. It is beyond doubt that intraspecific interactions have a far greater role in shaping these anatomies than interspecific ones, with the cranial ornaments of male giraffids, bovids, cervids and other taxa strongly influenced by sexual selection (Geist 1966; Bro-Jørgensen 2007; Knell et al. 2012). Their cranial structures are so strongly moulded by intraspecific adaptive pressures that they adopt sizes, shapes, textures and orientations that exclude them from effective predator defence (Estes 1991; Roberts 1996), instead becoming better suited to absorbing, catching and parrying the blows of rivals during physical intraspecific contests (Geist 1966; Packer 1983; Bro-Jørgensen 2007). They do not evolve these morphologies randomly, either, but change in response to specific fighting strategies and environmental circumstances. Horned female bovids may also engage in fights with other individuals (both male and female) of their species for resources, but their lessened behavioural emphasis on these bouts means their horns remain less developed than those of males — the significance of this is yet another area of discussion among zoologists.
So, having just thoroughly complicated this seemingly simple topic to a great extent, let’s bring this discussion back to dinosaurs and the assumption that horned dinosaurs wielded their horns like swords against dragon-esque theropods. While models of ceratopsids defending themselves with their horns are undoubtedly validated by the behaviour of some living species, a case can be made that we’ve overstated the importance of horns in predator defence among living animals and, by extension, dinosaurs. The message from the modern day is that horn-like structures can and might be used against predators, but that this behaviour is by no means ubiquitous. It may not even be that common, according to some researchers. It seems that intraspecific selection is more than sufficient to explain most horn-like structures among living species and that predatory influences, if present at all, are relatively minor for most species. We can’t know how much of this insight can be transferred over to dinosaurs, but if ceratopsid facial anatomy was being shaped by intraspecific factors (and we think it was; see above), then we have to entertain all that this brings. This means, in addition to the traditional view of horned dinosaurs being effective foils of predatory theropods, we have to consider some other possibilities suggested by their modern analogues. These could include, for instance, that only some horned dinosaurs actively fought predators; that their retaliations against attacks may have been ineffectual; and that some species may have rarely, and maybe never, turned their horns against other species. And this door swings another way: we have sufficient data from living animals to stop thinking that horned or spiked dinosaurs were the most formidable prey species and that “defenceless” dinosaurs like hadrosaurs and sauropods would be pushovers for their lack of obvious weaponry. Determining which fossil animals are “the most dangerous” from their raw anatomy overlooks the huge impact of non-fossilisable factors that contribute to anti-predator responses, such as temperament, prey awareness, physiology, intelligence, behavioural plasticity and so on. It’s a disappointing limitation of the fossil record that we can investigate what dinosaurs and other extinct animals were capable of, but we’ll never know what they were truly like. Questions about "the most dangerous dinosaur" and similar fall into that void.
Megasuperhypertheropod Tyrannosaurus encounters the unarmed sauropod Alamosaurus. "They said it was defenceless! Defenceless!"
So, in sum, the take-home here isn’t that anti-predator roles for ceratopsid horns are a non-starter, but that the behaviours of living animals complicate this seemingly simple hypothesis. If intraspecific evolutionary pressures on horns and related structures operate mostly independently of predatory pressures today, that has to be our model for Deep Time as well. This opinion comes loaded with caveats, of course, the biggest one being that we’re in a shifting landscape as goes determining the exact roles of horn-like structures in living species; as this changes, so might our ideas on extinct animals. And there’s a lot more we could discuss, too. There’s the aforementioned data about ceratopsid cranial functionality, there’s that healed, T. rex-bitten Triceratops horn described by Happ (2008) that is taken by some as evidence of defensive horn use (I’m not sure I agree; there’s no way of knowing the exact circumstances under which that horn was bitten), there’s the bigger picture of armed dinosaur co-evolution with different theropod clades… but we have to end here. I’ll conclude by borrowing a line from Farlow and Dodson (1975) who succinctly put the ceratopsid anti-predator hypothesis where it should be almost fifty years ago with the mere use of italics: “the evolution of ceratopsian cranial morphology probably reflects diversification through species-specific compromises among various selective pressures… and possibly predator resistance”.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Bakker, R. T. (1986). The Dinosaur Heresies. William Morrow and Company. Inc.
Bro‐Jørgensen, J. (2007). The intensity of sexual selection predicts weapon size in male bovids. Evolution, 61(6), 1316-1326.
Caro, T. M., Graham, C. M., Stoner, C. J., & Flores, M. M. (2003). Correlates of horn and antler shape in bovids and cervids. Behavioral Ecology and Sociobiology, 55, 32-41.
Chimes, L. C., Beytell, P., Muntifering, J. R., Kötting, B., & Neville, V. (2022). Effects of dehorning on population productivity in four Namibia sub-populations of black rhinoceros (Diceros bicornis bicornis). European Journal of Wildlife Research, 68(5), 58.
Colbert, E. H. (1948). Evolution of the horned dinosaurs. Evolution, 145-163.
D’Anastasio, R., Cilli, J., Bacchia, F., Fanti, F., Gobbo, G., & Capasso, L. (2022). Histological and chemical diagnosis of a combat lesion in Triceratops. Scientific Reports, 12(1), 3941.
Estes, R. D. (1991). The significance of horns and other male secondary sexual characters in female bovids. Applied Animal Behaviour Science, 29(1-4), 403-451.
Farke, A. A. (2004). Horn use in Triceratops (Dinosauria: Ceratopsidae): testing behavioral hypotheses using scale models. Palaeontologia Electronica, 7(1), 10p.
Farke, A. A., Wolff, E. D., & Tanke, D. H. (2009). Evidence of combat in Triceratops. PLoS One, 4(1), e4252.
Farlow, J. O., & Dodson, P. (1975). The behavioral significance of frill and horn morphology in ceratopsian dinosaurs. Evolution, 353-361.
Geist, V. (1966). The evolution of horn-like organs. Behaviour, 27(1-2), 175-214.
Geist, V. (1999). Deer of the World. Swan Hill Press, Shrewsbury.
Gerstenhaber, C., & Knapp, A. (2022). Sexual selection leads to positive allometry but not sexual dimorphism in the expression of horn shape in the blue wildebeest, Connochaetes taurinus. BMC Ecology and Evolution, 22(1), 107.
Happ, J. (2008). An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops. In Larson P. & Carpenter, K. Tyrannosaurus rex the Tyrant king, Indiana University Press. p. 355-370.
Horner, J. R., & Goodwin, M. B. (2006). Major cranial changes during Triceratops ontogeny. Proceedings of the Royal Society B: Biological Sciences, 273(1602), 2757-2761.
Horner, J. R., & Goodwin, M. B. (2008). Ontogeny of cranial epi-ossifications in Triceratops. Journal of Vertebrate Paleontology, 28(1), 134-144.
Knell, R. J., Naish, D., Tomkins, J. L., & Hone, D. W. (2013). Sexual selection in prehistoric animals: detection and implications. Trends in ecology & evolution, 28(1), 38-47.
Kruuk, H. (1972). The Spotted Hyena. A Study of Predation and Social Behavior. University of Chicago Press.
Mallon, J. C. (2017). Recognizing sexual dimorphism in the fossil record: lessons from nonavian dinosaurs. Paleobiology, 43(3), 495-507.
Matthew, W. D. (1915). Dinosaurs with Special Reference to the American Museum: Collections (No. 5). American museum of natural history.
Metz, M. C., Emlen, D. J., Stahler, D. R., MacNulty, D. R., Smith, D. W., & Hebblewhite, M. (2018). Predation shapes the evolutionary traits of cervid weapons. Nature Ecology & Evolution, 2(10), 1619-1625.
Osborn, H. F. (1917). The origin and evolution of life: on the theory of action, reaction and interaction of energy. C. Scribner's sons.
Packer, C. (1983). Sexual dimorphism: the horns of African antelopes. Science, 221(4616), 1191-1193.
Paul, G. S. (1988). Predatory dinosaurs of the world: a complete illustrated guide. Simon & Schuster.
Paul, G. S. (2008). The extreme lifestyles and habits of the gigantic tyrannosaurid superpredators of the Late Cretaceous of North America and Asia. In Larson P. & Carpenter, K. Tyrannosaurus rex the Tyrant king, Indiana University Press. p. 307-354.
Roberts, S. C. (1996). The evolution of hornedness in female ruminants. Behaviour, 133(5-6), 399-442.
Sampson, S. D., Ryan, M. J., & Tanke, D. H. (1997). Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and behavioral implications. Zoological Journal of the Linnean Society, 121(3), 293-337.
Schaller, G. B. (1972). The Serengeti Lion: A Study of Predator-Prey Relations. University of Chicago Press.
Spassov, N. B. (1979). Sexual selection and the evolution of horn-like structures of ceratopsian dinosaurs. Paleontol. Stratigr. Lithol, 11, 37-48.
Stankowich, T., & Caro, T. (2009). Evolution of weaponry in female bovids. Proceedings of the Royal Society B: Biological Sciences, 276(1677), 4329-4334.
A juvenile Edmontosaurus disappears into the enormous, lipped and gummy mouth of Tyrannosaurus. Those of us in the palaeoart community are used to seeing lips on dinosaurs now, but neither the lipped or lipless hypothesis has been given a thorough seeing-to in peer-reviewed literature yet. Until, that is, today. This is the PR art for our new paper, Cullen et al. 2023, that dives into the question of lips for theropod dinosaurs.
If you follow developments in palaeoart to any level of detail, you can’t have missed “the Lips Debate”: the controversy surrounding the application of extra-oral tissues (essentially various kinds of lips and cheeks) to extinct animals. This discussion has touched on virtually all fossil vertebrates at some time or another, but the presence or absence of lips on dinosaurs has, predictably, been the major focus for most palaeoartists. Owing to their general popularity, the lipped-or-not status of the predatory theropod dinosaurs has drawn a particularly large amount of attention. This debate is now so well covered online that its basic tenets will be familiar to many: the question of whether extant dinosaur relatives offer misleading insights for facial reconstruction; the importance of tooth size and angle to dental “sheathability”; the similarities and differences of jaw bone morphology between crocodylians, lizards and theropods, and so on.
But for all of this online visibility, the question of dinosaur lips and cheeks has received only a little attention from dinosaur researchers. A number of conference abstracts have been presented in this area, but only a handful of these studies have been pushed through peer review to become fully published scientific papers (e.g. Galton 1973; Ford 1997; Knoll 2008; Keillor 2013; Nabavizadeh 2020). It’s largely been dinosaur artists, mostly writing for blogs (such as this, this and this), social media posts or specialist books and magazines (e.g. Witton 2018; Paul 2019), that have provided the bulk of recent conversation on this issue. Whatever merits these discussions have (and there are some very fine, commendable assessments out there), the lack of detailed, authoritative scientific studies has allowed trends in dinosaur facial reconstruction to be shaped by popular culture, palaeoart memes and the opinions of influential palaeoartists more than conventional science. This means that, however comfortable we are with our opinions on dinosaur mouth appearance, this question would benefit from more study, more data and more insight from experts in reptile facial anatomy.
A visual review of where we are with restoring theropod mouths, from Cullen et al. 2023. Do you prefer your Tyrannosaurus without lips (B and C), or with lips (D and E)? And far more importantly, which of these is better supported by fossil data?
To that end, today is a good day. I’m part of an international team of researchers publishing a major new paper in Science dedicated to the question of whether theropod dinosaurs possessed lips. Led by Thomas M. Cullen and receiving contributions from Derek W. Larson, Diane Scott, Tea Maho, Kirstin S. Brink, David C. Evans and Robert Reisz, this is the end product of a long-running investigation into theropod faces that was first initiated by Robert 11 years ago. I was invited onto the project at the end of 2020 to create some artwork of lipped and lipless Tyrannosaurus (above) and, once onboard, I’m pleased to say I was able to help contextualise and interpret our data alongside providing some pretty pictures. But the real hard graft of the research was performed by others on the authorship team, so they deserve full credit for the nitty-gritty science and methodological concepts. They also taught me a lot about reptile jaws, teeth and oral soft tissues, so I’m in their debt for this experience.
As you’ll have guessed from the PR artwork that greeted you above, we conclude that yes, theropods almost certainly had lips. As evidence of this, we present multiple lines of evidence that all point to lizard-like scaly tissues covering predatory dinosaur teeth, and perhaps even other types of lizard-like oral tissues as well. Our work can be divided into four independent investigations that collectively support our assertions.
Jaw and tooth form
Firstly, we make some broad-brush comparisons between the tooth orientation and jaw bone morphology of lizards, crocodylians and theropods, some of which will be familiar to those who’ve followed the Lips Debate so far. We note that theropods, early croc-line archosaurs and lizards are similar in having low numbers of jaw bone foramina* distributed along their oral margins, as well as vertically-aligned teeth. Extant crocodylians, in contrast, have splaying teeth and hundreds of evenly-distributed foramina across their skull bones, the functions of which are more specialised than the lip-nourishing jaw openings of lizards.
*If you're new to all this, foramina are small holes in bones that typically house nerve tissues or blood vessels, but can also record other structures, like outgrowths of air sacs.
Reptile jawbone surface textures and foramina distribution compared, from Cullen et al. (2023). I think this image speaks for itself: the arrangement of theropod jaw foramina (those holes along the jawline) is far more lizard-like than croc-like.
Some of these observations are not novel as the significance of jaw bone foramina to the theropod lip question has long been recognised (e.g. Bakker 1986; Morhardt 2009; Keilor 2013; Barker et al. 2017; Carr et al. 2017). Our resurrection of this point is, in part, a response to Thomas Carr and colleagues' 2017 paper on Daspletosaurus horneri, which favourably compared tyrannosaur jaw surfaces to those of crocodylians. We don’t think they’re actually much alike at all, especially in foramina distribution, so disagree with that assessment. Carr et al. (2017) also assumed that the thick, immobile facial anatomy of living archosaurs — crocs and birds — was ancestral to their entire group, including non-bird theropods. We question this too. Like theropods, the jaw surface properties and tooth orientations of early croc-line archosaurs recall those of lizards more than modern crocodylians, probably reflecting a different soft-tissue configuration. We thus agree with the increasingly evidenced view that the faces of living archosaurs are specialisations suited to very particular lifestyles and that the dinosaur extant phylogenetic bracket is of limited use for inferring their facial anatomy.
Enamel, hydration, and tooth wear
Secondly, we discuss the damage and wear inflicted on permanently exposed teeth, a conversation that is mostly about enamel hydration. Enamel is one of the hardest tissues that animals can synthesise and is thus highly resistant to damage, but its resilience is dependent on moisture. Hydrated enamel is more plastic, and thus more resistant to abrasion, than dehydrated enamel, which is brittle and prone to cracking and breaking under strain. To that end, teeth emerging from oral margins tend to be more damaged and worn than those kept within a moist, sealed mouth. This difference can be seen with the naked eye but is particularly obvious under microscopic examination. In our paper, we show that the tips of alligator teeth are shorn off on their outward-facing, exposed surfaces, with both the enamel and several layers of dentine worn to a flattened edge. Tooth dehydration almost certainly factors into crocodylians frequently suffering from broken and cracked dentition, and they have to replace their teeth regularly (something like 45-50 times in a lifetime — Grigg and Kirchner 2015) to maintain a set of fully functional jaws.
These observations give us a clear hypothesis regarding theropod oral tissues. If they were permanently exposed, theropod teeth should show, at minimum, similarly obliterated enamel and dentine layers at their tips. The poster children of exposed dinosaur teeth, adult tyrannosaurids, are especially relevant here as they replaced their teeth at a very slow, sometimes even biennial rate (Erickson 1996). What’s more, they engaged in particularly violent, tooth-on-bone feeding strategies. So, if any theropods are going to have knackered, abraded teeth, it’s tyrannosaurids.
Detailed comparisons of tyrannosaurid (upper row) and crocodylian (lower row) tooth wear. Note how the Daspletosaurus tooth, despite being over 500 days old, is intact despite dinosaurs teeth having particularly thin enamel layers. The alligator tooth tip, by contrast, has not only lost the enamel coating on its outer surface, but also several layers of underlying dentine. A fully intact, enamel-covered erupting alligator tooth is shown in panel H to show that these are features of wear, not the original tooth condition. From Cullen et al. (2023).
But extracting a fully grown, c. 510-day-old tooth from a Daspletosaurus maxilla revealed a tooth tip in great condition. Both the inner and outer surface retained their relatively thin enamel covering and the only minor damage found was on its medial, inward-facing surface, possibly as a result of occasional tooth-on-tooth contact. This confirms what has generally been observed for theropod dental wear in other studies and conflicts with what we'd expect from a dehydrated, perpetually exposed tooth. We take this as evidence of theropod dentition being maintained in a moist, well-hydrated setting, and being located within a lipped mouth is realistically the only way this might be achieved.
Too big to sheath?
We also address the possibility that some theropods had teeth that were simply too big to cover with lips (e.g. Ford 1997). Our focus here is not on the widely known, but still surprisingly prevalent issue of artists and researchers not accounting for tooth slippage in fossil specimens**. Rather, we focus on realistic estimates of tooth crown height when they are fully socketed in theropod mouths. We calculated a ratio of tooth crown height to skull length for 37 theropod specimens and compared them with the same metric in 40 varanids, the monitor lizards. Varanids are, of course, well-known for possessing large, theropod-like teeth, as well as copious amounts of lip and gum tissue.
**This is the decay of tooth-anchoring ligaments resulting in teeth sliding somewhat from their sockets, preserving them at longer lengths than they held in life.
Tooth size: theropods vs varanids. It turns out that both groups have similarly-sized teeth relative to skull length and, while this doesn't directly tell us if theropods had lips, it shows that their dentition was of a size that we know can be sheathed by extra-oral tissues today. From Cullen et al. (2023).
Plotting these data showed that theropod and monitor teeth are about the same size for their skull lengths and even increase in proportion at the same approximate rate. But the winner of the biggest tooth contest wasn’t something like T. rex: it was the varanids. Some monitor species, like the crocodile monitor Varanus salvadorii, have almost cartoonishly-large dentition. From this, we suggest that theropods did not need unprecedentedly big lips to cover their mouths and the largest theropods wouldn't look, in terms of lip proportions, very different to something like a komodo dragon. Indeed, we note that monitors are able to cover their teeth with the same basic configuration of labial and gingival tissues across a 12-fold size difference. The discrepancy between the largest monitor skull and our largest theropods is only half that: 6-fold. So if lizard lips and gums can scale 12 times over without substantial anatomical deviation, perhaps they could stretch to cover the teeth of much bigger animals without much change, too? Whether we've realised it or not, a lot of us already evidently believe this is possible, given the abundance of lippy, monitor-esque mosasaur reconstructions.
Sealing the deal
These are the points we cover in the main paper but, this being an article in Science, it’s essential to also check out the supplementary files for additional discussion and context. Therein we raise another point that resulted from our efforts at reconstructing a scientifically-informed illustration of a lipless Tyrannosaurus: it’s really, really hard, maybe even impossible, to seal at least some theropod mouths without lips. Forming an oral seal, even if it’s just by pressing lipless jaws against one another, is important to avoiding dehydration as well as maintaining basic oral health and hygiene. We could not, however, find a way to reconstruct T. rex jaws without leaving a gap behind their maxillary teeth. I actually pushed our lipless reconstruction (Fig. 1B in the paper) a little beyond what I think is reasonable and we're still left with a small opening.
We are not the first people to ponder this issue, and dinosaur literature has contrasting views on how far theropods could close their jaws. Some authors propose that theropod mandibles could be pulled way up into the cavity of the upper jaw and have even identified landmarks for the resting position of the lower teeth (below). These include depressions in the walls and roof of the upper oral chamber that seem suited to act as socket-like structures for receipt of the lower dentition (e.g. Molnar 1991; Ford 1997; Currie 2003; Hendrickx et al. 2014). Others, most notably Tyler Keillor (2013) in his excellent book chapter on restoring the face of the “Jane” Tyrannosaurus, have questioned this idea on grounds that theropod mandibles can’t close so tightly without literally bashing into problems.
Examples of landmarks suggested to record the resting poses of theropod lower jaws. Maxillary wall sockets are depressions in the internal wall of theropod oral cavities, and some theropods are also preserved with round depressions in the roofs of their mouths. Neither are universal features of all theropods, however. Images from Osborn (1912), Lü et al. (2014) and Cullen et al. (2023).
Reconstructing the face of T. rex for our paper saw us agreeing with Tyler's conclusions. At a certain point of mouth closure, theropod lower jaws collide with bones under the eye socket (specifically, the ectopterygoid) so that further adduction either requires the jaws to literally crush themselves shut, or else the bones of the posterior skull act as a hinge, swinging the jaw tip into the mouth but dislocating the jaw joint. Ford (1997) proposed that a notch in the ectopterygoid accommodated the closed lower jaw during mouth closure but we don’t think this is plausible. Theropod ectopterygoids can be complex shapes and yes, some have regions that superficially look like they could nestle the lower jaw, but these were almost certainly filled by deep jaw muscles in life (e.g. Gignac and Erickson 2017). There are, of course, theropod skulls preserved with their jaws tight shut in the fossil record but we have to be careful assuming these represent in vivo conditions, given how routine processes of decay and fossilisation can pull and crush carcasses into unnatural configurations.
Inspired by this, we devote some discussion to how theropods posed their closed jaws in life. Beyond ruling out impossible, jaw-busting configurations, it's difficult to know exactly how tightly theropods held their mouths, but this is something for artists to consider. X-rays and scans of lizard carcasses show that their jaws are far from clenched shut when their mouths are closed and, in some species, their upper and lower dentition barely overlaps. If we go "full lizard" with our theropod reconstructions, where we apply minimal overlap of the upper and lower toothrows, their lips would have been deep and their snouts much taller than we’re used to. We play about a little with this visually in the paper and I was struck at the blocky, chunky cranial profile of our lizard-like, loose-mouthed T. rex (below).
Some of our experimentation with theropod mouth postures. The "crush closed" pose is almost certainly impossible, but it's hard to say how relaxed theropod resting gapes may have been held. If modern lipped reptiles are anything to go by, they may have been held far more "open" than we're used to. Modified from Cullen et al. (2023).
Conclusion: theropod jaws only make sense if they had lips
Putting all this together, our investigations of jaw structure, tooth size, tooth wear and jaw closing all point to the same inference: theropod jaws don’t make much anatomical or functional sense without lips of some kind. To validate the alternative lipless model, we have to engage in a lot of special pleading and scientific weaselling. Exposed theropod teeth would have to be unprecedentedly resistant to wear; all our understanding of jaw structure and foramina distribution correlating with oral soft tissues would have to be wrong, and theropods would need to be unique in not bothering to create oral seals. If we're being good scientists, we can’t currently say that theropods definitely had lips, drop the microphone and walk off stage, but I think we've made it far more challenging for anyone to legitimately object to the lipped theropod hypothesis. Time will tell on that front.
In addition to substantiating the lipped hypothesis of theropod appearance, our hope is that our paper may establish some lines of inquiry for the oral tissues of other extinct animals. Many of the most extreme dentitions to ever evolve belong to fossil taxa, after all, and theropods are far from the only species with uncertain facial appearances. What of nimravids, gorgonopsians, uintatheres, or Thylacosmilus? And what, for that matter, of the superficially crocodile-like spinosaurids and other weird theropods — were they lipped or not? If our ideas hold water, they provide a relatively straightforward way of deducing whether the teeth of these animals were held within oral tissues.
We can't, of course, finish without some brief notes on the life appearance of lipped theropods. We address this a little in our supplementary information and conclude that lepidosaurs, the lizards and tuataras, are the best modern analogue for theropod lips and gums. This is, admittedly, a “best of a bad situation” recommendation because there are plenty of differences between theropod and lepidosaur jaws that preclude total confidence in their comparison, but we only have so many extant reptile groups to choose from and lepidosaurs are, on the whole, morphologically closer to theropods in areas we think are influential on labial and gingival tissues.
What did lipped theropods actually look like? Lizard mouths have a lot more soft-tissue surrounding their teeth (jaw cross sections show a komodo dragon, B, and alligator, C), and this might be something we need to bring into our theropod artwork. The gaping T. rex shown here is outfitted not only with large lips, but also a conservative 25% of its tooth height covered with gums. From Cullen et al. (2023, supplementary data).
This being the case, our model for theropod mouths is that they were sealed by non-muscular*** lips covered with scales or — to hedge our bets a little more — whatever epidermal covering was present on the side of the snout. Lepidosaurs show variation in lip size, with most having generous upper lips but some having thinner lower lips than others. This variation continues to their gums. Lepdiosaur gingivae are more voluminous than those of mammals and crocodylians and generally cover at least 20-25% or so of tooth crown height. This is why lizard teeth aren’t always that conspicuous in their open mouths. Varanids take these enlarged gingivae to an extreme, hiding almost all of their formidable teeth with enormous gums. We currently don’t have much insight into where theropods sat within this range. Paul (2019), independently of our study, advocates for full monitor-like conditions for theropods, and this might be possible, but we can't rule out smaller gums or, indeed, a unique theropodan take on oral soft-tissues at this time. We propose, however, that since extant lipped reptiles have at least 25% of their tooth crown heights covered with gums, we should apply that to theropods, too. We’ve explored this in our paper and PR art with that gummy, lippy T. rex shown above.
***It’s not strictly true that lizards have no muscles around their mouths. Some agamids have muscles that move their lips or flaps of mouth-adjacent skin for communicative purposes. I'm no expert on these structures, but I think it’s fair to assume that they are specialisations of their respective lineages, not the remnant of a once ubiquitous, clade-wide lepidosaur ability.
And I think we’ll leave it there for now. There’s more to say on the tyrannosaur restorations we created for the paper with their small eyes and some of their skin details, but at least some of that discussion needs to wait for developments in other research I’m involved in (I'm so, so sorry, Dave). For now, I’ll thank my coauthors once again for inviting me onto such a great project, and I’ll leave you with this handy infographic summary of our research, which you can share around the internet to instigate discussion of dinosaur life appearance wherever you may be. The QR code in the corner will take you directly to the paper so, if you or anyone you encounter wants to know exactly what we have to say about theropod lips, you can always find it from this image.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Bakker, R. T. (1986). The dinosaur heresies: new theories unlocking the mystery of the dinosaurs and their extinction. William Morrow.
Barker, C. T., Naish, D., Newham, E., Katsamenis, O. L., & Dyke, G. (2017). Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii. Scientific Reports, 7(1), 3749.
Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports, 7(1), 1-11.
Cullen, T. M., Larson, D. W., Witton, M. P., Scott, D. Maho, T. Brink, K. S., Evans, D. C. and Reisz, R. (2023). Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology. Science, 379, 1348-1352.
Currie, P. J. (2003). Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada.
Ford, T. L., 1997, Did Theropods have Lizard Lips? Southwest Paleontological Symposium – Proceedings, 1997, 65-78.
Galton, P. M. (1973). The cheeks of ornithischian dinosaurs. Lethaia, 6(1), 67-89.
Gignac, P. M., & Erickson, G. M. (2017). The biomechanics behind extreme osteophagy in Tyrannosaurus rex. Scientific Reports, 7(1), 2012.
Grigg, G., & Kirshner, D. (2015). Biology and Evolution of Crocodylians. Csiro Publishing.
Hendrickx, C., & Mateus, O. (2014). Torvosaurus gurneyi n. sp., the largest terrestrial predator from Europe, and a proposed terminology of the maxilla anatomy in nonavian theropods. PloS one, 9(3), e88905.
Keillor, T. M. (2013). Jane, in the flesh. In: J. M. Parrish, R. E. Molnar, P. J. Currie, E. B. Koppelhus, (eds.). Tyrannosaurid Paleobiology. Indiana University Press.
Knoll, F. (2008). Buccal soft anatomy in Lesothosaurus (Dinosauria: Ornithischia). Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 248(3), 355-364.
Lü, J., Yi, L., Brusatte, S. L., Yang, L., Li, H., & Chen, L. (2014). A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nature communications, 5(1), 3788.
Molnar, R. E. (1991). The cranial morpholgy of Tyrannosaurus rex. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie, 217, 137-176.
Morhardt, A. C. (2009). Dinosaur smiles: Do the texture and morphology of the premaxilla, maxilla, and dentary bones of sauropsids provide osteological correlates for inferring extra-oral structures reliably in dinosaurs?. Western Illinois University.
Nabavizadeh, A. (2020). New reconstruction of cranial musculature in ornithischian dinosaurs: implications for feeding mechanisms and buccal anatomy. The Anatomical Record, 303(2), 347-362.
Osborn, H. F. (1912). Crania of Tyrannosaurus and Allosaurus; Integument of the iguanodont dinosaur Trachodon. Memoirs of the AMNH; new ser., v. 1, pt. 1-2.
Paul, G. S. (2019). Non-ornithischian dinosaurs probably had lips. Here’s why. Prehistoric Times 127, 44-49.
Witton, M. P. (2018). The Palaeoartists' Handbook. Crowood Press.
Finally: a modern book devoted to everyone's favourite mid-20th Century Czech palaeoartist, Zdeněk Burian. It's big, it's expensive, and it's amazing. For palaeoart fans, it's an essential purchase.
As much as I’m a fan of 19th century palaeoart, the 20th Century is where our efforts to reconstruct fossil organisms really take off and stretch their wings. In its early decades, Charles R. Knight led a charge to reinvigorate and re-legitimise palaeoart after enthusiasm for illustrating the life appearance of fossil organisms cooled in the late 19th century. Later, the likes of Gregory S. Paul and Jay Matternes transformed how palaeoart would be executed and valued by scientists. And between these trailblazers lies the career of another palaeoartistic giant, an individual who took Knight’s mantle as the most important palaeoartist of his day and carried it almost until his death in 1981: Zdeněk Burian.
Born of Czech descent in Austria-Hungary in 1905, Burian began his palaeoartistic career in the mid-1930s. Much has been said about Burian over the years and his status as a master of palaeoart is unquestioned. Whether we see him as tag-teaming with Knight to define the second half of a “classic palaeoart” era (Witton 2018) or as the lead of another phase, termed “modern palaeoart" by Manucci and Romano (2023), Burian defined the look of prehistoric animals for many of us born in the 20th century. Already a well-established illustrator before being headhunted for high-profile palaeoart projects, Burian took on his first prehistoric subjects just as Knight's career began winding down, and his prodigious artistic ability and eye for natural history made the young Burian a great and intuitive palaeoartist from the start. Heavy promotion from his collaborators pushed his work into the same space of international recognition once held by his American forerunner. Influencing or simply outright copied by countless other illustrators, Burian remained a leading palaeoartist until the final days of his career. The torch was only passed during the late 1970s when a new generation, armed with new science and palaeoartistic philosophy, changed the rules of restoring extinct animals and — for better or worse, depending on your perspective — introduced new aesthetics and values into the discipline (Lescaze 2017; Witton 2018; Manucci and Romano 2023; Müller et al. 2023).
There are lots of famous images of sauropods from the mid-20th century, but few are as iconic as Burian's 1941 Brachiosaurus. Yes, this scene is impossible but no, that doesn't affect the quality or influence of this image one bit. Borrowed from Albatros Media.
Given this pedigree, it’s mysterious that Burian’s prehistoric artworks have not been written about as much as those by Knight, Waterhouse Hawkins, or even recent artists like Jay Matternes, at least in the English-speaking world (e.g. Czerkas and Glut 1982; Paul 2000a; Bramwell and Peck 2008; Milner 2012; Carrano and Johnson 2015; Witton and Michel 2022). Take a look at any palaeoart bookshelf and the lack of a decent Burian-themed collection is an obvious and major hole. Until recently, filling this space was only possible by collecting second-hand copies of Burian’s collaborations with Josef Augusta, Zdeněk Spinar and Vratislav Mazák. But with the vintage of these texts reaching or well surpassing 50 years, they are increasingly difficult to find or are prohibitively expensive — rarer Burian texts retail for hundreds of pounds in the UK.
It’s into this void that Albatros Media is injecting their new tome Prehistoric World of Zdeněk Burian Volume 1: From the Creation of the Earth to the Extinction of the Dinosaurs(or, in its original Czech, Pravěký svět Zdeňka Buriana - Kniha 1: Od vzniku Země po zánik dinosaurů). Written and compiled by Ondřej Müller, Bořivoj Záruba, Martin Košťák and Rostislav Walica, it represents the first of an ambitious three-part series compiling the totality of Burian’s palaeontological paintings, illustrations and sketches, both familiar and obscure, and many never published before. Available through the Albatros Media website and also select retailers (see below), this first volume may prove to be the most popular as, along with his Precambrian and Palaeozoic artworks, it contains all of Burian’s pre-Cenozoic dinosaur pieces. These, of course, include many unquestionably iconic pieces of palaeoart: water-bound Brachiosaurus (above), blue-green Archaeopteryx, rearing Tarbosaurus and so on. The book is written in Czech and there are reportedly no plans for a full English translation, but English speakers are catered for with some in-book caption details as well as an introductory essay on the Albatros website derived from two important chapters.
Undoubtedly, splitting Prehistoric World of Zdeněk Burian into three books is the only way to document Burian’s hyper-prolific output. Despite containing just a subset of his prehistoric-themed artwork, this first volume is enormous. Covers measuring 24 x 33 cm sandwich 600 pages of heavy, high-quality paper into a spine almost 6 cm thick. Containing over 400 images, many of which are displayed at large size, the whole package is just a few grams shy of 4 kg. I’ve been tempted to try a patented Tetrapod Zoology Podcast-style table drop to hear the noise of the thud, but I worry about what might happen… to the table. It’s remarkable that another two books, presumably of similar size, will be necessary to document Burian’s palaeontological artwork given that this was not the main focus of his career: he continued to provide illustrations for other books and these, not his extinct animals, form the bulk of his portfolio. I look forward to seeing all three of the palaeo volumes together, even if I am a little concerned about what 12 kg of new books might do to my already creaking bookshelves.
I'm not kidding about that nigh-on 4 kg weight. In future, we'll be able to identify Burian fans by their especially large, well-toned biceps.
Such size carries a cost: €99 for European buyers (excluding VAT and shipping) and $130 in the US. That aims Prehistoric World of Zdeněk Burian Volume 1 squarely at palaeoart diehards and suggests a price tag north of €/$300 for all three volumes. As steep as that might initially seem, it’s actually a bargain compared to the cost of collecting Burian’s art through secondhand book markets. Moreover, with the majority of artworks being rescanned or re-photographed for this project, as well as the inclusion of 135 pieces from private collections, this will be the definitive Burian palaeoart archive for many years to come. Price-conscious buyers may be pleased to know that the illustrations are grouped by geological age rather than by date of production (see below) so you can selectively buy the volume with your favourite time periods or taxa if desired. I suspect there is scope for a coffee table book that compiles the most famous Burian pieces into a more affordable product, but that’s another project, for another time. For now, palaeoart and Burian fans are in for a treat: every penny of that cover cost has gone into making Prehistoric World of Zdeněk Burian Volume 1 an exceptional piece of work.
Given that I can’t read Czech, I can only comment briefly on the text via the online introduction and two translated chapters. These cover the outlay of the book, Burian’s palaeoartistic career and his artistic process. Müller et al. (2023) explain their overarching (but not religiously followed) geochronological and systematic arrangement as the best manner to catalogue Burian’s work because he returned to the same subjects, and sometimes even the same compositions, again and again. Displaying his art in order of production may have given a greater perception of Burian’s developing talent and career, but would probably have been confusing and repetitive to read. Instead, we are presented with everything Burian created for different time periods — the complete Burian Archaen, the complete Burian Cambrian etc. — within which artworks are ordered taxonomically. Only here, at this granular level, does Burian’s professional history affect image order, with his oldest takes on a given topic presented first.
The iconic Burian 1970 Tarbosaurus in Müller et al. (2023, left) compared to its appearance in Z. V. Spinar’s 1972 Life Before Man (right). Even in this wobbly phone photo, the difference in quality is obvious.
This approach makes for an intuitive way to tour Burian’s portfolio and, with a turn of the page, we can compare different eras of his illustrations of the same subjects. We see that his style somewhat tightened over time, the looser qualities of his earliest work turning into more exacting brushstrokes and finer detail by the 1950s. He also revised and repainted many illustrations in light of new science. The latter is an overlooked aspect of Burian’s palaeoartistry because it can, owing to its volume, ubiquitousness and the sometimes subtle alterations he made to older paintings, seem homogenous and largely unchanging when viewed in isolation. But when presented together, we see that Burian’s portfolio is arguably one of the most scientifically transformative of any palaeoartist. In spanning the 1930s to the earliest 1980s, his career captured wholly different, often strongly contrasting palaeontological perspectives. Sometimes viewed as following the “modernist consensus” of dinosaur science (Paul 2000b), Müller et al. (2023) argue that Burian pushed boundaries in dinosaur reconstruction where he could, such as by depicting mobile-wristed and charging ceratopsids as early as the 1940s and 1950s, as well as embracing Bakkerian dinosaur form in the 1970s. I wonder what Burian, who would have been in his 70s when working on these “new look” dinosaurs, thought of producing such radically different takes on subjects after 40 years of creating “traditional”, now old-school reconstructions.
Burian's 1976 Barosaurus, an example of the "giraffoid Barosaurus" meme started by Bob Bakker. We can criticise this painting for being derivative of Bakker's original but, for 1976, this was still cutting-edge stuff: compare this to the Brachiosaurus above, drawn decades earlier. As noted by Müller et al. (2023), Burian's paintings sometimes borrowed elements from other artists because he lacked access to specimens himself, and had to work pretty much exclusively from secondary sources of paleontological information. Image from Albatros Media.
Müller et al. (2023) give some fascinating insights and observations on Burian’s career and artwork, both of scientific and cultural nature. Among the most interesting and, to my knowledge, novel, concerns how Burian’s palaeoart served as propaganda. The prominence of his art in the West makes it easy to forget that Burian lived and worked on the eastern side of the Iron Curtain, and Müller et al. (2023) argue that he ultimately benefitted from socialist rule despite the hard times faced by artists in Czechoslovakia during the mid-20th century. A communist coup saw private publishers nationalised in the late 1940s and this put many illustrators out of work, potentially including Burian. But his palaeoartistic collaborations with Josef Augusta were officially supported and encouraged by the new regime because of their embodiment of Darwinism, a philosophy embraced by Marxist-Leninist outlooks. Somewhat ironically, Burian’s own conviction on evolution was not so firm; his take on life's development was open-minded and pantheistic in nature. Regardless, he profited from the capacity of palaeoart to promote ideals important to a socialist government and his illustrator career was not only rescued from ruin, but destined for opportunities among lauded authorities and individuals. Müller et al. (2023) do not go so far as to compare Burian's career with those of other famous palaeoartists (at least, not the in the texts I can read) but it’s hard not to see his professional life as a political mirror of Knight’s, who found his own fame via support from the independently wealthy, thoroughly capitalist benefactors of American museums.
Müller et al. (2023) further comment on Burian’s approach and attitude to art, and their view is a little more complex and nuanced than that offered recently by Zoe Lescaze (2017). Lescaze posits that a troubled upbringing, a strained relationship with his father, a penniless independence at the age of 15 and the near-constant threat of war and political upheaval contributed to what she perceives as a savage, brutal art style. She sees his paintings as being populated by “sinister dinosaurs”, “thuggish Neanderthals” and other “monsters charged with an almost carnal corporeality”. Müller et al. (2023) agree with this to a point but caution against “foreign theorists” stereotyping Burian by interpreting his work as being the product of a politically troubled state. While also finding melancholy in Burian’s lonely, sad-looking dinosaurs and his fragmented hominid family groups, they marry the tinge of sadness and isolation in Burian’s paintings with the more pragmatic, aspirational and upbeat sides of his character. It's noted that his compositions were often shaped by his forensic attention to scientific detail, his desire to teach through his paintings, and an “uncomplicated and playful” worldview that was captivated by nature. This multifaceted interpretation of Burian’s seems more likely to me than the barely-contained savagery suggested by Lescaze, especially because, for all his dozens of scenes of animal confrontations, physical violence is not a feature characterising his artwork. You can thumb all the way through Prehistoric World of Zdeněk Burian Volume 1 and find virtually no violence or gore.
Burian's moody Monoclonius. If you asked me to place this in palaeoart history I would have suggested the 1970s or 1980s for its somewhat Stoutian qualities. But no, this is from 1948: Burian was taking palaeoart to strange new places well before the modern push for greater artistic styles and approaches. Another swipe from Albatros Media.
Of course, one does not really buy a compilation of Zdeněk Burian palaeoart just for the text: what about the presentation of the artwork itself? On this most critical matter, Prehistoric World of Zdeněk Burian Volume 1 truly excels. It seems that no expense has been spared in documenting everything Burian created on prehistoric subjects, from preparatory sketches and drawings to paintings and illustrations in all sorts of styles and media. All are reproduced at good sizes and in excellent detail. Short of looking at the actual artworks themselves, it’s hard to imagine getting a better view of his creations. Unsurprisingly, the visualisation is far superior to that of older books and yes, you definitely want these versions for their improved colour balances and detail even if you already have a copy of something like Spinar’s Life Before Man or Augusta’s Prehistoric Animals on your shelf (see direct comparison, above). Look closely and you’ll note that even the surface texture of Burian’s oil pieces is evident. The printed page can’t replicate the actual three-dimensional quality of his paintings with their thick paint ridges or their picked and threadbare, canvas-exposed regions, but you certainly get a sense of their presence. Special mention should be made of the multitude of plans, notes, roughouts and sketches that feature alongside finished pieces. We rarely or only tokenistically document these peripheral but enlightening pieces in our palaeoart literature, so their inclusion is very welcome here.
My only slight gripe with the images concerns layout, as many are stretched across two pages in a way that loses details in the page binding. This is a concern for any book but, in this 6 cm deep monster, the page gutter is a deep chasm, especially if the book is not laid flat on a table. But to mention this is nit-picking of the highest order and I’m ultimately moaning about a design no-win scenario: if seam issues were avoided by consigning images to one page only, I’d probably be asking for bigger pictures.
Preview pages of Prehistoric World of Zdeněk Burian Volume 1 compiled from Albatros Media's website. Note the diversity of images: ink drawings, sketches, annotated illustrations and full paintings.
Indeed, there’s only one real issue with Prehistoric World of Zdeněk Burian Volume 1: it’s too good. In this first book alone, Müller et al. (2023) have set the standard for documenting our palaeoart masters so high that other efforts look underwhelming by comparison. If volumes 2 and 3 follow suit, the Prehistoric World of Zdeněk Burian series will be the gold standard for documenting any palaeoartist, and the fact Ondřej Müller, Bořivoj Záruba, Martin Košťák and Rostislav Walica have achieved this for someone as prolific as Burian is nothing short of a triumph. The amount of care and investment that’s gone into this project is extraordinary and it will take an especially dedicated team to create a palaeoart compilation for another artist that can stand shoulder-to-shoulder with it. And that, for the record, is the new goal: I’m not sure any collection will ever surpass it. Yes, it really is that good.
Prehistoric World of Zdeněk Burian Volume 1: From the Creation of the Earth to the Extinction of the Dinosaurs (Pravěký svět Zdeňka Buriana - Kniha 1: Od vzniku Země po zánik dinosaurů) is out now. It is internationally available from Albatros Media as well as select retailers and distributors, including Donald M. Grant in the US and NHBS in the UK. European buyers can also email burian@albatrosmedia.cz for more information.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
Bramwell, V., & Peck, R. M. (2008). All in the Bones: a biography of Benjamin Waterhouse Hawkins. Academy of Natural Sciences
Carrano, M. T. & Johnson, K. R. (2015). Visions of Lost Worlds: The Paleo Art of Jay Matternes. Smithsonian Books.
Czerkas, S. M., & Glut, D. F. (1982). Dinosaurs, mammoths, and cavemen: the art of Charles R. Knight. Dutton Adult.
Lescaze, Z. (2017). Paleoart: Visions of the prehistoric past.
Milner, R. (2012). Charles R. Knight: the artist who saw through time. Abrams.
Müller, O, Záruba, B, Košťák, M & Walica, R. Rostislav. (2023). Pravěký svět Zdeňka Buriana - Kniha 1: Od vzniku Země po zánik dinosaurů. Albatros Media.
Paul, G. S. (2000a). The art of Charles R. Knight. In Paul, G. S. (ed). The Scientific American Book of Dinosaurs. Macmillan. 113-118 pp.
Paul, G. S. (2000b). A quick history of dinosaur art. In Paul, G. S. (ed). The Scientific American Book of Dinosaurs. Macmillan. 107-112 pp.
Witton, M. P. (2018). The Palaeoartist’s Handbook: Reconstructing Extinct Animals in Art. Crowood Press.
Witton, M. P. & Michel, E. (2022). The art and science of the Crystal Palace Dinosaurs. Crowood Press.

![[page image]](http://images.library.wisc.edu/HistSciTech/EFacs/CompAnat/0009/M/PLATEVI.jpg)
![[page image]](http://images.library.wisc.edu/HistSciTech/EFacs/CompAnat/0012/M/PLATEIX.jpg)
![[page image]](http://images.library.wisc.edu/HistSciTech/EFacs/CompAnat/0008/M/PLATEV.jpg)




















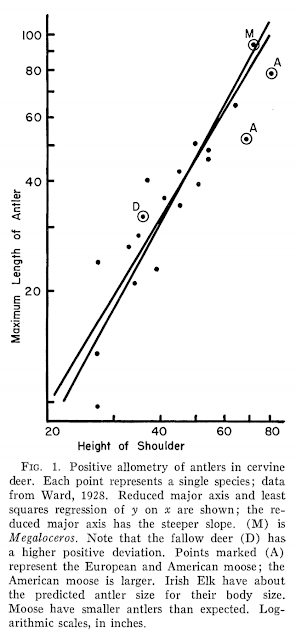















































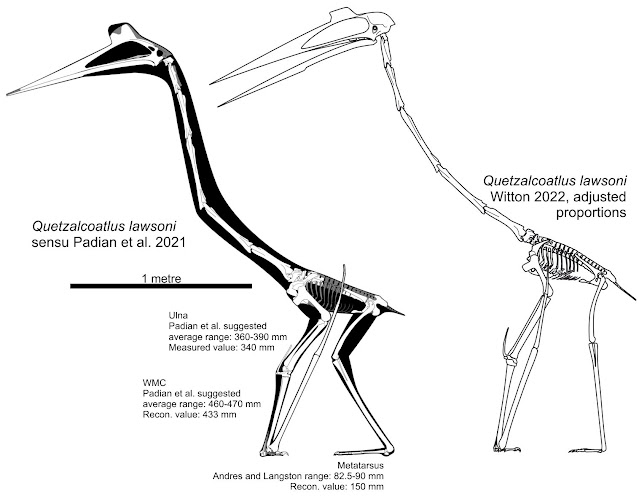
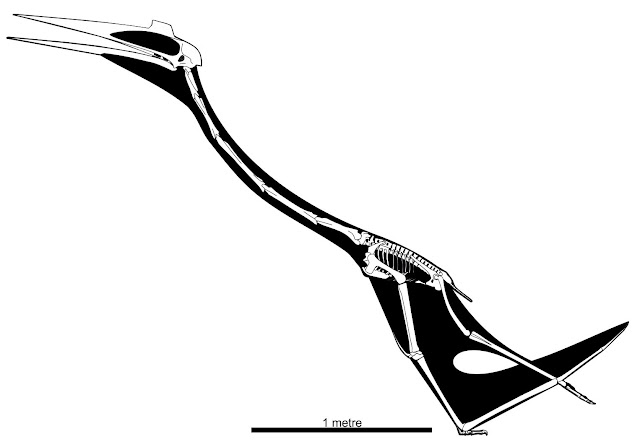
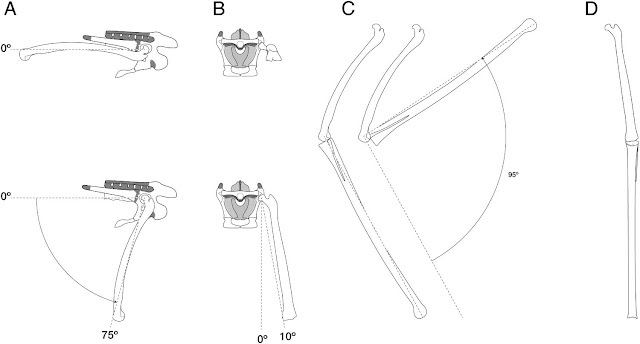
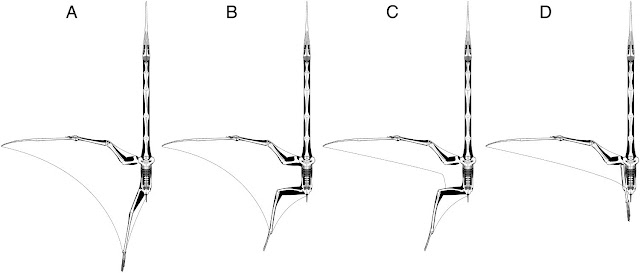
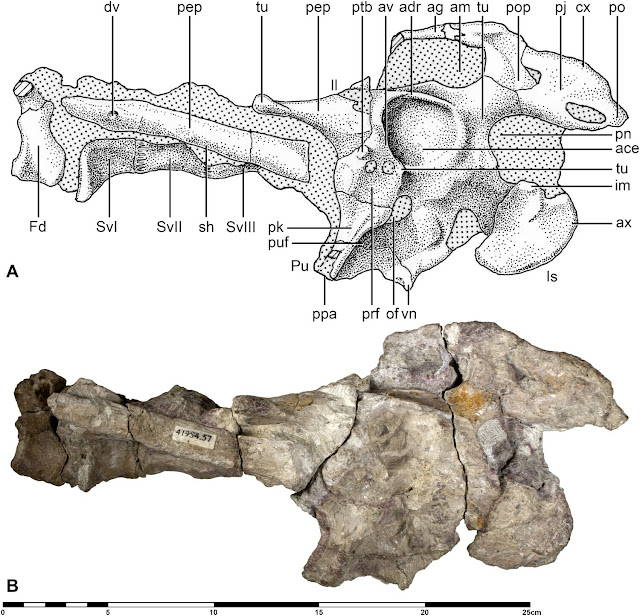
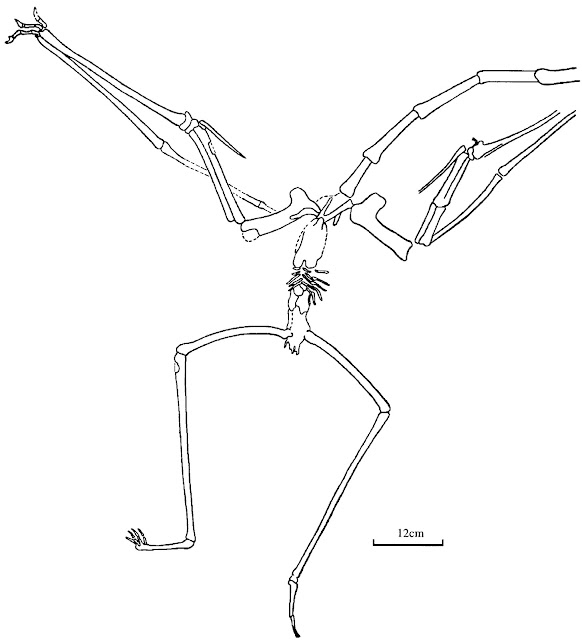
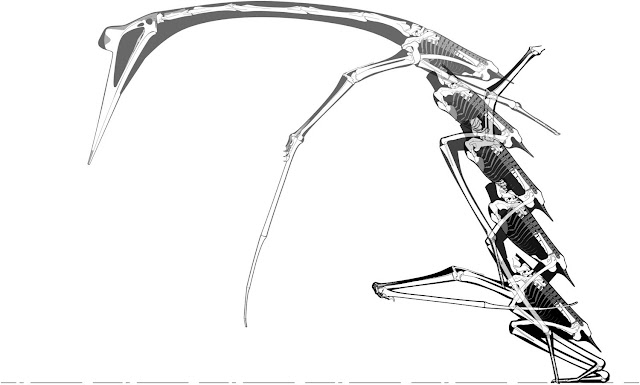




































.jpg)
.jpg)



























.jpg)

















