The story of how dinosaurs were resurrected for cinema in the early 20th century tends to focus on the visual components of these technical feats: the 2D animation and stop-motion technology that made long-extinct reptiles move again. But alongside making dinosaurs walk and run, cinematic dinosaurs also gave them a voice, imagining their snarls, barks, screeches and roars. Perhaps the first film to do this (or certainly the first mainstream, widely-seen film, at any rate) was 1933’s King Kong, where a charging Stegosaurus might take the title of first vocalising dinosaur in cinematic history:
The same film would give us plenty of other prehistoric animal action and noise, but by far the most famous is surely the snarling, screeching Tyrannosaurus:
Over the last century, cinematic dinosaur appearance and the technologies used to depict them have changed dramatically but one thing has remained the same: dinosaurs are as loud and noisy as ever. We’ve even seen the development of conventions and tropes around what dinosaurs sounded like. Sauropods are often given haunting, humpback whale-like songs and dromaeosaurs are frequently assigned aggressive, high-pitched snorts, snarls and crackles. And, of course, big theropods — especially Tyrannosaurus — invariably have deep, bellowing roars. It’s no exaggeration to say that the Jurassic Park Tyrannosaurus roar is as iconic and recognisable as its now-famous (maybe infamous?) design.
The desire to depict talkative, raucous dinosaurs has long transcended media capable of conveying sound. Even movie dinosaurs were noisy and boisterous before we had the technology to make them truly roar on screen. The 1925 silent film The Lost World features plenty of roaring, snarling and bellowing stop-motion dinosaurs framed by director Harry O. Hoyt in dramatic close-up. Even in silence, the intent of these shots is obvious, and we simply have to imagine their vocalisations ourselves. Fantasia’s famous 1940 Rite of Spring sequence performed a similar trick for artistic reasons, juxtaposing a roaring Tyrannosaurus against booming segments of Igor Stravinsky’s famous composition. Static, traditional palaeoart also has a strong emphasis on animal vocalisations. Peruse any gallery of prehistoric animal restorations (such as this, at my new website!) and we inevitably find heaps of artworks showing grunting, chirping, screaming dinosaurs. Viewed objectively, it is a little strange that we focus so much on this behaviour in our artwork. Why don’t we render more non-acoustic behaviours that are arguably better suited to a totally visual medium? And furthermore, why draw so much attention to an aspect of dinosaur behaviour we don't know much about? Subconsciously, we just can’t get away from the call — pun not intended — of depicting extinct animals vocalising.
 |
| An Archaeopteryx siemensii perches on driftwood, opens its mouth and calls out... what, exactly? We palaeoartists can't resist drawing animals posed mid-vocalisation, despite our lack of knowledge about extinct animal sound production. |
Clearly, we've collectively decided that prehistoric reptiles were vocal, noisy species, and this is understandable. It is, after all, what we experience around us today. Our world is full of singing, calling birds and barking, bellowing mammals. Whales sing, lions roar, and frogs croak. It stands to reason, then, that dinosaurs would be just as vociferous, and that a Jurassic or Cretaceous dawn would be full of strange, wondrous hoots, bellows, chirrups and songs that we can only imagine. I am, of course, leading up to a weighty “however”. What if our assumption of noisy, vociferous dinosaurs is simply... wrong?
This is, of course, a very strong accusation, especially because we can say very little definitively about dinosaur vocalisation owing to our lack of fossilised dinosaur throat tissues and vocal organs. The preservation of such anatomies among Mesozoic dinosaurs is not impossible, these having been found in Vegavis iaai, a Mesozoic bird that lived in Antarctica 69-66 million years ago (Clarke et al. 2016); but this remains an exceptional occurrence: Vegavis is the only Mesozoic dinosaur known with preserved vocalisation anatomy. We can, however, use fossils and data from extant reptiles and birds to make predictions about dinosaur vocal ability, and several researchers have attempted this (e.g. Weishampel 1981; Senter 2008; Brazaitis and Watanabe 2011; Clarke et al. 2016; Reide et al. 2016). Among the more famous examples of such works is Phil Senter’s 2008 Voices of the past: a review of Paleozoic and Mesozoic animal sounds, a synthesis of what we know of sound production among ancient animals. In his section on birds and their ancestors, Senter makes the bold suggestion that non-avian dinosaurs may have been — yikes — entirely non-vocal (Senter 2008). In other words, this posits that dinosaurs may have not only been much quieter than their pop culture counterparts, but actually reliant on non-vocal acoustics when they wanted to communicate audibly. This notion — which I’m calling the "silent dinosaur hypothesis” — gained a fair bit of discussion online when first published and still crops up in modern conversations about dinosaur behaviour. But how does it hold up over a decade on, and did it ever have a sound basis to begin with?
 |
| Non-vocal (or, at least, closed-mouth) interaction between male and female Ceratosaurus nasicornis. Were dinosaurs limited to posturing and other means of display for their communication with one another? Some hypotheses suggest so. |
To explore this further, it will help to outline what non-vocal animal acoustics are. We animals make noise all the time simply by existing and going about our lives. These ‘passive’ noises are classed as non-vocal acoustics. They include sounds that come from acts like breathing, forcing air around your throat tissues, and hitting or rubbing body parts against each other or external objects. Many animal species exploit these phenomena to make deliberate, structured sounds for communication. A hiss, for example, is little more than forcefully expelling air through our throats and mouths. A snort is much the same, except using our noses. We can also purposefully slap or rub body parts together or against the ground, water or vegetation to create loud noises. Some species have developed special anatomy purely to create non-vocal sounds, with the most obvious example being rattlesnake tails. Non-vocal acoustics are everywhere once we start noticing them, and Senter (2008) argued that they may have been the only sounds made by dinosaurs. It's incontrovertible that these are the only noises we can be confident that dinosaurs made because they can be generated regardless of vocal capability. Whatever other noises dinosaurs created, we know that they could hiss, snort, stamp their feet and so on, and living diapsids show that such behaviours are used as communication strategies among extant dinosaur relatives. Non-vocal acoustics are also perfectly compatible with the large noses and crests that may have acted as resonating chambers in certain dinosaurs, too (e.g. Weishempel 1981; Witmer and Ridgely 2009). We know, for instance, that some non-vocal snake species use resonating cavities in their throats to turn hisses into growls (Young 1991; see an example here of the slightly terrifying noises from of a king cobra).
So, yes, non-vocal acoustics make a lot of sense for dinosaurs — documentary makers, take note. But Senter's (2008) accusation is that dinosaurs could only make non-vocal sounds, and that requires us to consider 'true' vocalisations: the sounds animals make by forcing air through their vocal organs. Mammals, amphibians and non-avian reptiles use a larynx for this purpose, while birds have their own, unique voice organ: the syrinx. It’s the evolution of this structure that prompted Senter’s suggestion of non-vocal dinosaurs. Unlike the larynx, which is situated at the top of the throat, the syrinx is located at the base of the trachea where the airway forks into the lungs. It also works in a different way to a larynx. Rather than passing air over vocal folds, the syrinx generates sound from the airway walls themselves. Rushing air from the lungs flutters these membranes and associated cartilages in a manner that produces sound, and the location of the syrinx at the fork where the trachea becomes a pair of bronchial tubes allows for especially complex vocalisations: each bronchial component can vibrate asymmetrically, making two sounds at once. To keep their airways open, avian syrinxes are reinforced with well-mineralised cartilaginous rings. In modern birds, it seems that syrinxes can also only function with assistance from a clavicular air sac (Senter 2008), although experiments indicate vocalisation without this structure may be possible (Clarke et al. 2016).
 |
| Thanks to fossils of Vegavis, we can be confident that extinct duck and goose relatives like Conflicto antarcticus — a Palaeogene species from Antarctica shown here — were capable of making honking, goose-like sounds. But we have very little direct evidence for the sort of noises more rootward dinosaurs were able to make. |
The reinforced structure of the syrinx and its possible association with an air sac means that, unlike larynxes, they have some geologically detectable elements. In theory, this allows us to gauge roughly when, and in which lineages, they evolved even if fossil syrinxes themselves are rare. We can search for evidence of the clavicular air sac pneumatising the bones of the pectoral girdle and forelimb, as well as fossils of those reinforcing, mineralised rings holding the syrinx open. Although soft-tissue in nature, these structures are found in Cenozoic bird fossils (Clarke et al. 2016), so they evidently have decent enough fossilisation potential in the right circumstances.
The results of such searches have come back without much to report, however. Even in well-preserved Mesozoic dinosaurs, we find no consistent evidence for clavicular air sacs outside of the ornithothoracines (the group of avialans that includes enantiornithines and crown birds, but see Senter 2008; Wedel 2009 for a few exceptions) and not a single mineralised airway has been discovered in a non-avian dinosaur (Senter 2008; Clarke et al. 2016). This suggests that the syrinx was developed very late in dinosaur evolution, perhaps not even being present in feathered, otherwise extremely-bird-like dinosaurs (Clarke et al. 2016; Kingsley et al. 2018). We should not assume, of course, that the avian syrinx sprang into existence fully-formed — surely it had to develop via intermediary ‘proto-syrinx’ structures first (Kingsley et al. 2018) — but we don’t know what that structure was nor what features might evidence its existence. With our present dataset, all we can say is that the avian syrinx as we know it probably wasn’t present in most non-bird dinosaurs. It follows that if dinosaurs did not have a syrinx, they were probably incapable of making the rich, complex noises of modern birds.
And this is where things get especially interesting. OK, so dinosaurs weren't singing like passerines, but most reptiles have a larynx, and we can be pretty certain that dinosaurs did too. So Senter must be wrong, right? Dinosaurs merely vocalised like modern reptiles: case closed. Well, not necessarily, because we don't know if the dinosaur larynx was functional. Many lizard larynxes lack vocal folds and thus cannot vocalise, and opinions differ on whether their vocal abilities were independently lost from a vocal reptilian ancestor (e.g. Kingsley et al. 2018) or convergently gained from a historically silent one (e.g. Russel and Bauer 2021). Furthermore, birds also have a larynx, but it's also non-functional. This leaves dinosaurs evolutionarily bracketed by crocodylians (with a functioning larynx) and birds (with a non-functioning larynx), creating ambiguity about the ancestral state of dinosaur vocal organs. The ancestral acoustic capabilities of other reptiles is thus very important to determining what the original state of archosaur vocalisation was. There are two possible models (Kingsley et al. 2018): perhaps archosaurs were ancestrally silent, with crocodylians and birds to developing functional vocal organs independently of one another; or they were vocal, with birds augmenting and/or replacing the larynx for an unknown reason late in dinosaur evolution. And this touches on another key question with bearing on dinosaur vocalisation: why did birds develop the syrinx at all? One possibility is that the syrinx evolved in response to having lost, or having never developed, a vocal organ in the first place (Kingsley et al. 2018), a scenario implying that at least some theropods, if not all dinosaurs, went through a silent phase in their evolutionary history. The bottom line is that there's still a lot to learn about the evolution of reptile vocalisation, and there are reasonable, entirely plausible models that align with Senter’s (2008) proposal that dinosaurs were non-vocal (below).
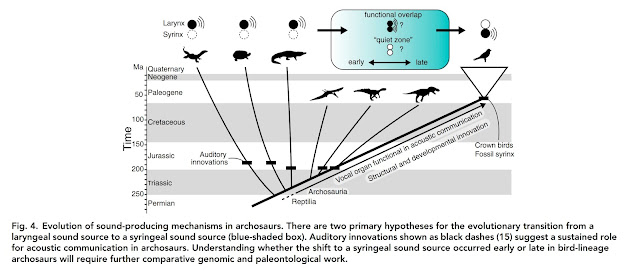 |
| A handy graphic showing two competing models of syrinx evolution, from Kingsley et al. 2018. This assumes that reptiles were ancestrally vocal, but this doesn't change considerations of syrinx evolution too much. Essentially, we have two options: birds evolved a syrinx alongside a functioning larynx, or the dinosaur larynx wasn't functional, and the syrinx evolved as a novel structure to exploit vocal communication. The latter model, of course, implies at least some non-vocal dinosaurs. |
These ideas are, of course, very difficult to test without appropriate fossil data. Senter (2008) noted some support from non-vocal lizards using visual communication instead of aural, thus placing extra significance on the often extravagant display structures of dinosaurs. Might all those crests, horns, frills, fancy scales and elaborate feathers have evolved because dinosaurs were essentially mute, primarily visual communicators (Senter 2008)? A counterargument to this is that lizards communicate visually without such crests, horns and so on, but the concept of some dinosaurs using display structures to compensate for a lack of vocal capability is still an interesting idea.
But before we get carried away with all this, we should note that the silent dinosaur hypothesis is not the only model of archosaur acoustic evolution on the table. A case can be made that, whatever weirdness was going on with syrinx evolution, dinosaurs were still capable of making laryngeal sounds. It has been noted that birds and crocodylians share several similar vocal behaviours that implies inheritance from a shared, vocal ancestor (e.g. Brazaitis and Watanabe 2011; Clarke et al. 2016) and some models of reptile evolution posit that all reptiles were ancestrally vocal, implying a functioning larynx in Dinosauria (Kingsley et al. 2018). Such concepts predict that dinosaurs vocalised at least in relation to matters of territory and courting, as well as to communicate between parents and offspring (Clarke et al. 2016). It’s difficult, of course, to know what specific sounds were made, and this isn't just because larynxes rarely fossilise: it's also because reptilian vocal anatomy is just not as well studied as that of birds and mammals (e.g. Rittenhouse et al. 1998; Reide et al. 2015; Russel and Bauer 2021). Recent work has shown that, although most reptile vocalisations are relatively simple compared to those of frogs, mammals and birds, there is a lot of variation in larynx structure across reptile species, and that their vocal tissues and acoustic capabilities can be very sophisticated, sometimes competing with mammals and birds in complexity (Brazaitis and Watanabe 2011; Reide et al. 2015; Russel and Bauer 2021). Among the most developed reptile vocal capabilities are those of crocodylians, which include a repertoire of behaviourally-specific hisses, grunts, bellows, snorts and chirps (Garrick et al. 1978), and those of geckoes, which use a range of single and repetitive chirps for advertising and alarm purposes (Russel and Bauer 2021). Perhaps, assuming dinosaurs did have functional larynxes, they made similar sounds.
We should clarify that the comparisons made by some researchers between reptilian and mammalian larynxes does not necessarily imply that dinosaurs vocalised like mammals. The throat tissues of reptiles and mammals are quite different in that reptiles can inflate their neck tissues with air from their lungs, whereas mammalian throat cartilage and muscles prohibit this action (Reide et al. 2016). This equips diapsids with a distinct mechanism for loud, deep sound production: closed-mouth vocalisation. By closing their mouths to prevent air escape and pumping air into their necks, reptiles and birds can create resonating chambers which allow for much deeper, lower-frequency vocalisations than could be achieved with a 'standard' open mouth call. We might intuitively think of crocodylians employing this behaviour to create loud, awesome bellows (especially the American alligator, which is the champion of crocodylian bellowing - see Garrick et al. 1978 and video above) but this tactic is not just used by big, exotic species: the cooing of pigeons and the ‘a-woo’ of eider ducks are also closed-mouth vocalisations. These acoustics have developed repeatedly throughout archosaur evolution and may have been practised by the dinosaur-crocodylian ancestor (Reide et al. 2016), so it seems reasonable to imagine this behaviour being used by dinosaurs making especially loud, deep and far-reaching noises. This may have been especially so among large species as, in birds at least, closed-mouth vocalisations have mostly evolved among bigger-bodied lineages (Reide et al. 2016). Predictions of archosaur voice evolution do not suggest that all dinosaur vocalisation would be closed-mouth (Reide et al. 2016), but those of us interested in depicting dinosaurs making their loudest, most intimidating noises should consider closed-mouth behaviours more likely than the usual stereotype of cat-like roaring (indeed, the roaring ability of Panthera species is associated with an unusual throat and laryngeal configuration (see Weissengruber et al. 2002), so we shouldn’t regard it as a ‘typical’ noise for any extinct animal to make, especially a reptile).
But we're getting a little off-topic now: we're here to talk about silent dinosaurs, not booming ones. So let's wrap things up. To summarise, there are a few take-homes here. The first is that the general assertion that we know nothing about dinosaur vocalisation isn’t really true: we certainly don’t know much, but we’re not entirely devoid of intelligent comment, either. A lot of the papers cited in this post are available online and are well-worth reading if you want to know more about the topics discussed above. The second is that the silent dinosaur hypothesis is far from a done-deal, but it has a more legitimacy than we might first expect. It's not, despite its unorthodoxy, a crazy idea and actually fits some interpretations of dinosaur vocal evolution, even if we can't really tell how right or wrong it is at the moment. There are huge caveats around any model of dinosaur vocal evolution, of course, the most important being that our models are so poorly informed by fossil data that one new discovery could turn everything we’ve predicted on its head. And that leads to a third main point: whatever ideas of dinosaur vocalisation we think are correct, we should appreciate that they’re not much more than personal preferences at the moment. But that’s fine, and it's even liberating for artists and filmmakers. This uncertainty gives us a huge playground for depicting dinosaur behaviour in ways we haven’t considered before. What does a non-vocal Tyrannosaurus do instead of roaring when it wants to look impressive? How do sauropods communicate without singing? What noises did Mesozoic birds make before they developed the syrinx? We don’t know, but it’s sure a heck of a lot of fun to think about.
Enjoyed this post? Support this blog for $1 a month and get free stuff!
This blog is sponsored throughPatreon, the site where you can help artists and authors make a living. If you enjoy my content, please consider donating as little as $1 a month to help fund my work and, in return, you'll get access to my exclusivePatreon content: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!
References
- Brazaitis, P., & Watanabe, M. E. (2011). Crocodilian behaviour: a window to dinosaur behaviour?. Historical Biology, 23(01), 73-90.
- Clarke, J. A., Chatterjee, S., Li, Z., Riede, T., Agnolin, F., Goller, F., ... & Novas, F. E. (2016). Fossil evidence of the avian vocal organ from the Mesozoic. Nature, 538(7626), 502-505.
- Garrick, L. D., Lang, J. W., & Herzog, H. A. (1978). Social signals of adult American alligators. Bulletin of the AMNH; v. 160, article 3.
- Kingsley, E. P., Eliason, C. M., Riede, T., Li, Z., Hiscock, T. W., Farnsworth, M., ... & Clarke, J. A. (2018). Identity and novelty in the avian syrinx. Proceedings of the National Academy of Sciences, 115(41), 10209-10217.
- Riede, T., Li, Z., Tokuda, I. T., & Farmer, C. G. (2015). Functional morphology of the Alligator mississippiensis larynx with implications for vocal production. The Journal of experimental biology, 218(7), 991-998.
- Riede, T., Eliason, C. M., Miller, E. H., Goller, F., & Clarke, J. A. (2016). Coos, booms, and hoots: The evolution of closed‐mouth vocal behavior in birds. Evolution, 70(8), 1734-1746.
- Russell, A. P., & Bauer, A. M. (2021). Vocalization by extant nonavian reptiles: a synthetic overview of phonation and the vocal apparatus. The Anatomical Record, 304(7), 1478-1528.
- Senter, P. (2008). Voices of the past: a review of Paleozoic and Mesozoic animal sounds. Historical Biology, 20(4), 255-287.
- Weishampel, D. B. (1981). Acoustic analyses of potential vocalization in lambeosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology, 7(2), 252-261.
- Weissengruber, G. E., Forstenpointner, G., Peters, G., Kübber‐Heiss, A., & Fitch, W. T. (2002). Hyoid apparatus and pharynx in the lion (Panthera leo), jaguar (Panthera onca), tiger (Panthera tigris), cheetah (Acinonyx jubatus) and domestic cat (Felis silvestris f. catus). Journal of anatomy, 201(3), 195-209.
- Witmer, L. M., & Ridgely, R. C. (2009). New insights into the brain, braincase, and ear region of tyrannosaurs (Dinosauria, Theropoda), with implications for sensory organization and behavior. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 292(9), 1266-1296.