 |
| A 2020 take on some dinosaur or another. I forget its name. This individual has recently gorged itself, resulting in a distended belly and sleepy demeanour. |
Unless you've been living under a rock for the last fortnight you cannot have escaped news on one of the most famous and controversial of all dinosaurs: Spinosaurus aegyptiacus. The appearance of Spinosaurus has once again transformed via the discovery of new fossils unearthed from the Late Cretaceous Kem Kem beds of Morocco: chiefly, a long paddle-like tail of superficially newt or crocodylian-like flavour. Keen interest in Spinosaurus, as well as a large National Geographic-led PR campaign for the new study, has seen social media awash with discussion about the new discovery, and illustrations of the latest in spinosaurine fashion have swamped online galleries since. As Chris Dipiazza eloquently explained on Twitter, it hasn't been the best two weeks if you aren't a Spinosaurus fan.
Okay fine I did the #Spinosaurus with it’s new paddle tail. Everyone’s debating on what the purpose of this wonderous tail was. Well I think it’s quite obvious, don’t you? We just observed it in real time. #scicomm#paleontology#theropodspic.twitter.com/wi8GjxKJnf— Christopher DiPiazza (@ChrisDPiazza) April 30, 2020
At the risk of numbing everyone further to Spinosaurus, I want to share some thoughts and reactions to this new research here. We've covered a few (but not all) of the twists and turns of Spinosaurus research in recent years (posts 1, 2, 3, 4) but, rather than simply writing another blog summary or popular rehashing of the new findings, I thought I'd write this from an artistic perspective, based on reading I conducted to produce my own take on "Spinosaurus 2020", shown above. As keen-eyed readers will note, I've not slavishly stuck to the same interpretations currently circulating the press circuit because - as we'll see - our takes on Spinosaurus are more complex than ever. Even with the tremendous amount of new data published on Spinosaurus in the last two decades, itremains the ultimate moving target for dinosaur palaeaoartists.
Spinosaurus 2020: where are we now?
Before we dive into this post, it makes sense to go beyond the recent Spinosaurus press coverage to look at what's in the new papers, as there's a lot more to them than what is being reported in the popular press. Needless to say, Spinosaurus has been an especially hot topic in dinosaur palaeontology since 2014 when Nizar Ibrahim and colleagues placed a newly discovered partial skeleton from the Moroccan Kem Kem beds at the core of a radical reinterpretation of a genuinely enigmatic animal. This was our introduction to Spinosaurus as a potentially short-legged semi-aquatic species, as well as proposals that spinosaurine material from across Northern Africa should be collected into one species, S. aegyptiacus, sinking several named taxa in the process (Ibrahim et al. 2014). It also proposed that the new partial skeleton should be the replacement exemplar specimen - the neotype - for S. aegyptiacus, after the original was destroyed in the Second World War to leave us with only Ernst Stromer's original descriptive work and photographs as records of its existence. Though widely publicised and catalysing a wave of public interest in Spinosaurus, the proposals of Ibrahim et al. (2014) proved controversial among academics. Numerous responses - some supportive, some critical - have been published by theropod researchers since.
 |
| Spinosaurus as illustrated by Ibrahim et al. 2014. Many aspects of this reconstruction have been questioned and scrutinised in the last six years, but new data suggests that this may be closer to the appearance and proportions of certain spinosaurines than many of us initially believed. |
Fast forward to today, and we've got not one, but two new papers by Nizar and colleagues that push discussions of all things Spinosaurus along significantly. The first is not Spinosaurus specific, but is an extensive monograph on the geology and palaeontology of the Kem Kem beds, now formally termed the Kem Kem Group (Ibrahim et al. 2020a). This is an important paper that brings some long-needed clarity and formality to details of Kem Kem stratigraphy and geology, including a new nomenclatural scheme to divide the Kem Kem into the Gara Sbaa and Douira formations. This is an important step for understanding the provenance of Kem Kem fossils which clarifies details of palaeoenvironments, relative ages of fossils, and comparisons with other fossil faunas (Ibrahim et al. 2020a). The entire fossil record of the Kem Kem Group is also reviewed, including a large discussion about Spinosaurus and its status as a Kem Kem theropod. Anyone interested in Spinosaurus and its world will need to check this paper out.
The second paper concerns additional material of the same 'neotype'* individual published in 2014, recovered from the same locality in more recent fieldwork (Ibrahim et al. 2020b). Among other finds, this includes a nearly complete tail that reinvents the appearance of Spinosaurus for the second time in six years. In addition to being short-limbed, it now seems that Spinosaurus had a deep, fin-like tail comprising narrow vertebrae with long and reclined neural spines and chevrons. This is interpreted as evidence of a swimming predatory ecology by the authors, it being argued that Spinosaurus could have swum like a crocodylian or newt to chase prey. The associated PR provides large amounts of media - videos, artwork etc. - showing Spinosaurus as a deep-diving species adapted to chasing large fish. This is not the first time Spinosaurus has been interpreted as a strong swimmer in recent years (e.g. Gimsa et al. 2016; Arden et al. 2019) but the recovery of a fin-like caudal skeleton adds a lot more weight to this argument.
*I'm going to refer to this specimen as the 'neotype' throughout this article for readability, as it's a catchier name than FSAC-KK 11888. That the proposal that FSAC-KK 11888 should be the Spinosaurus neotype remains controversial however, and will likely remain so until it's fully described and we can properly evaluate its similarity to Spinosaurus. I don't have a horse in this race but, for what it's worth, FSAC-KK 11888 looks like a member of Spinosaurus to me, although it has several differences from S. aegyptiacus that require investigation.
 |
| Artistic speculations that Spinosaurus may have borne some sort of tail fin have been common since the 2014 reinterpretation of the genus. Here's my finned version from 2016. |
Both papers also provide comments in defence of Spinosaurus palaeobiology as proposed by Ibrahim et al. (2014). Criticism of their work included doubts about the authenticity and scaling of the neotype skeleton (Evers et al. 2015; Henderson 2018); its biomechanical feasibility as a swimming animal (Henderson 2018); the appropriateness of collating widely-dispersed and anatomically-distinguished North African spinosaurine material into one species (a question with particular reference to the overall number of spinosaurines in the Kem Kem) (Evers et al. 2015; Hone and Holtz 2015; Hendrickx et al. 2016; Maganuco and Dal Sasso 2018; Lakin and Longrich 2019); the suitability of the proposed Spinosaurus neotype (Evers et al. 2015; Maganuco and Dal Sasso 2018); and the general ecology of spinosaurines (Hone and Holtz 2015, 2019; Henderson 2018). The new data presented by Ibrahim et al. (2020a, b) addresses some of these concerns to an extent that some criticism - as we'll explore below - can probably be laid to rest. However, the enhanced debate around all things Spinosaurus means that these new papers have arrived in a much busier and more heated academic realm than their 2014 counterpart, and initial impressionsfrom key players in spinosaurine research imply conversations will remain ongoing about aspects of lifestyle and taxonomy. For artists, this complicates our view of what Spinosaurus and other spinosaurines may have looked like, as well as what we can show it doing. While contributing important primary data on Spinosaurus, we have to remember that these new papers represent one interpretation of the appearance and lifestyle of a most unusual dinosaur in an increasingly busy academic debate, and that the ball is still in play.
Body plan and proportions
With that set up, it's time to dig into some art-relevant details. Firstly, I think Ibrahim et al. (2020b) adds a lot more confidence to the proposed strange proportions of Spinosaurus. The authenticity and scaling of the neotype have been questioned on grounds that it was collected, purchased and excavated by different people at different times (Evers et al. 2015; Henderson 2018), but the recovery the new tail and other elements in the same site as the pelvic, hindlimb and torso material, as well as their concordant proportions, suggests that all these remains were genuinely associated and likely belong to one individual (Ibrahim et al. 2020b). There is no evidence of other species in the bonebed and many broken bones of the neotype have now been reunited with once-missing pieces. Their histology and inferred growth stage are also matching. Courtesy of a quarry map illustration, we have a good idea of how these elements were associated in the field and how they relate to the material published in 2014.
 |
| Quarry map of the neotype locality and skeletal reconstruction of Spinosaurus, from Ibrahim et al. (2020b). Note the large area in which bones were found, the absence of non-spinosaurine bones, and the absence of bone duplicates: this is good evidence of the neotype representing a single individual, no matter how peculiar its proportions are. Known elements of the neotype are shaded in the skeletal, with different colours reflecting different field seasons and quarry locations. Scale bar represents 1 m. |
With these data, and the fact that another spinosaurine specimen (Stromer's "Spinosaurus B") shows the same short-limbed morphology (Ibrahim et al. 2014), I think we can be fairly confident that at least some spinosaurines really were long-bodied, short-legged creatures with a body plan basically akin to that outlined by Ibrahim et al. (2014, 2020b). I know some folks are still holding out for data proving that the pelvis and hindlimbs belong with the vertebral column, but I think the burden of proof has shifted in light of these new data. Why aren't these legs associated with the body, given what we now know about the taphonomy of the site? A common question online is how much bearing the new tail has on other spinosaurids. We have sufficient skeletal remains of baryonychine spinosaurids (e.g. Baryonyx, Suchomimus) to suggest that they weren't fin-tailed, but the tails of spinosaurine spinosaurids aren't well known. The dorsal and caudal vertebrae of Ichthyovenator compare well with Spinosaurus, however (Allain et al. 2012), and it may have sported similar tail anatomy.
Posture and balance
Within the supplementary data of Ibrahim et al. (2020b) is a discussion of Spinosaurus mass and centre of gravity based on the (estimated) 11 m long neotype individual. Using a digital model and varying takes on tissue density, a mass of 3,219-4,173 kg was predicted and the centre of gravity was found to be just over one femur-length from the pelvic limb joint. This is fractionally more posterior than modelled in the 2014 model, if not quite as close to the pelvis as predicted by Henderson (2018). The cause of this shift is the larger tail and, although subtle, this difference has forced a reassessment of one of the most controversial aspects of the 2014 study: the presentation of Spinosaurus as a quadruped. Ibrahim et al. (2020b) now favour a facultative, rather than obligate, quadrupedal gait for terrestrial locomotion.
For artists, this means we can be a little more comfortable posing Spinosaurus as a biped, and I wonder if further work will substantiate bipedal poses further. Elsewhere in the supplementary data, Ibrahim et al. (2020b) suggest that the volume of restored tail musculature is conservative, and it stands to reason that models with more substantive tail volumes will pull the centre of gravity rearwards. Moreover, I wonder if the restored neck bulk is a little on the thick side, making the model more front-heavy. Among the neotype elements are long cervical ribs which, assuming typical tetrapod neck anatomy, could indicate displacement of some ventral neck muscles towards the torso (Taylor and Wedel 2013). Given that Spinosaurus already seems to have had a longish, low neck skeleton, displacing some of the neck muscle fraction posteriorly could have made for a relatively slender neck that would lighten the front end. If Spinosaurus also walked a little more upright than a typical theropod - using poses proposedby Andrea Cau, say - it might have avoided quadrupedality altogether.
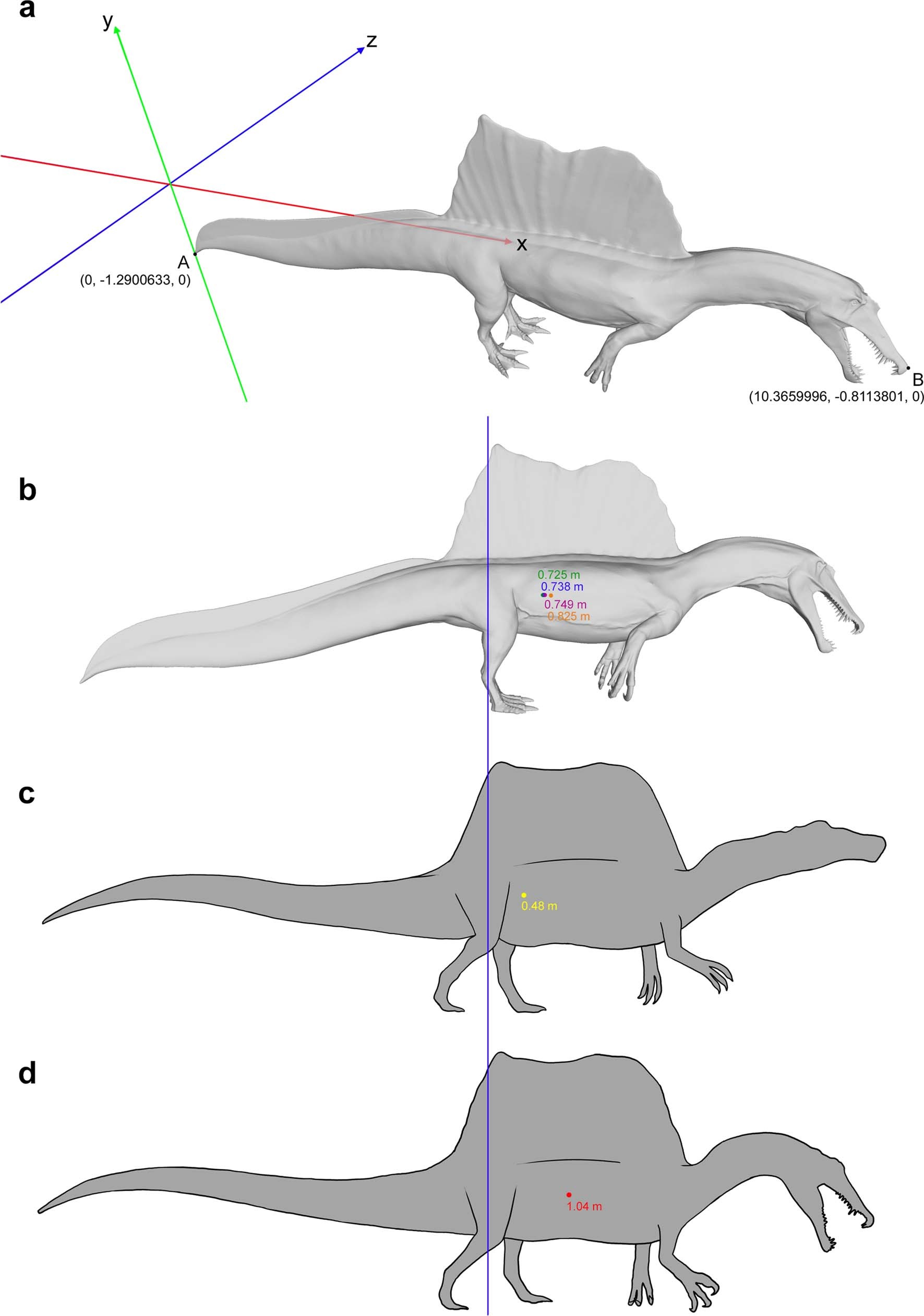 |
| Centre of mass estimates from Ibrahim et al. (2020b), compared to that of Henderson 2018 (C) and Ibrahim et al. 2014 (D). |
I'm aware that some people feel that the legs of the neotype specimen are too slender to support the weight of Spinosaurus on land. The predicted 3 - 4-tonne masses of the neotype individual are relatively lightweight compared to theropods of similar length (>10 m theropods in the dataset of Benson et al. 2014 mass at 6-7 tonnes, for instance) and the hindlimbs would have to be held pretty straight to clear the animal from the ground (see illustrations, above). If so, the hindlimbs might have been loaded more like columns and imparted greater support than a traditionally bent theropod limb. Using hindlimb measurements from Ibrahim et al. (2014), I ran some very basic calculations on the strength of the neotype femur and found it critically weak against bending: it would fail when loaded with less than one 4-tonne body weight. When loaded as a column, however, it could take multiple 4-tonne masses. These calculations were very basic and ignore a lot of the nuance associated with theropod femoral posture but, if basically accurate, they suggest that the hindlimbs were strong enough to support Spinosaurus on land without help from weight-bearing forelimbs. I won't share the full details of these sums here as this post is already very long, but I can produce a follow-up article if it's of interest. Furthermore, while the hindlimbs themselves are small, there is evidence that aspects of their musculature - such as the caudofemoralis (a powerful hindlimb retractor) - were not reduced. In occupying much of the top half of the femur, the fourth trochanter of the neotype Spinosaurus femur is proportionate to the rest of the body (see for yourself in the 2014 image above, panel I, label 'ft') and suggests that the legs were capable of propelling their owner forward with suitable force, perhaps without propulsive assistance from the forelimb.
A topic I'm going to avoid here is the swimming posture of Spinosaurus, as this is an area that warrants further investigation before anything concrete can be said. I feel that the digital floating experiments with the 2014 Spinosaurus reconstruction by Don Henderson (2018) presented several worthy criticisms of Spinosaurus as an underwater swimmer, including its inability to sink due to pneumatised skeletal components (though some bones of Spinosaurus were pachyostosic (Ibrahim et al. 2014), its skull, neck and dorsal vertebrae were not) and the elevated centre of mass created by the tall, dense sail. A caveat about this study is that Spinosaurus had a relatively wider torso than was factored into the floating model, which would likely impact placement of the centre of mass and thus stability. We shouldn't dismiss Don's work because we assume this will correct the tipping issue, however: we need to see this investigated. We also have to consider the impact a wider torso would have on the suggested 'unsinkable' nature of Spinosaurus, as a wider torso will increase the lung volume fraction and impact buoyancy. For the time being we perhaps need to recognise that the body plan of Spinosaurus, even with its new tail, is entirely unlike any swimming animals alive today and that it's challenging to know how it functioned in water. Our science on this unusual dinosaur is in its infancy, and forming robust ideas about its swimming pose and capability is going to take time.
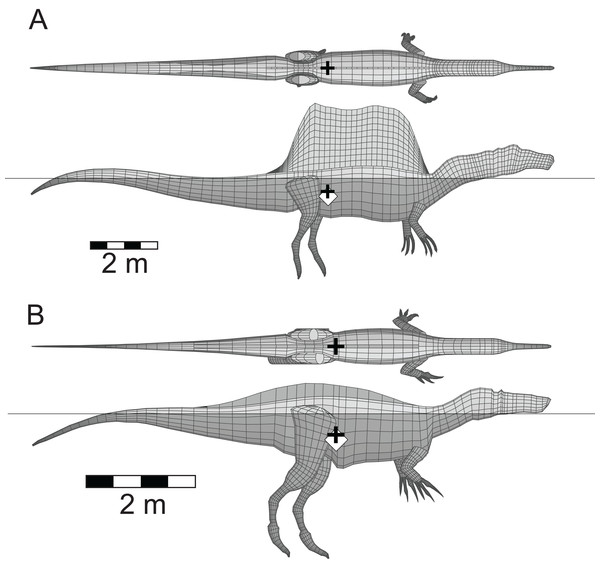 |
| Floating spinosaurids from Henderson (2018). One of the take-homes from Don's work is that Spinosaurus did not have an unusual floating posture among theropods, and that theropods were, in general, capable of floating with their heads well clear of the water to breathe. This questions whether features of the Spinosaurus skull linked to aquatic lifestyles - like the position of the eyes and nose - were specific adaptations to aquatic lifestyles. |
Sail shape
One area where I'm less certain about the proportions of our new Spinosaurus reconstruction is the shape of the torso sail. Reconstructing the sail shape of Spinosaurus has always been difficult because the original S. aegyptiacus vertebrae were already not in great shape before Allied bombs blew them to pieces. As shown in Stomer's 1915 plates, the Spinosaurus neural spines were mostly disassociated from their centra; some were broken or deformed at their tips; and their arrangement within the vertebral series was not clear, even to those who saw them in person (Smith et al. 2006). Accordingly, several ideas about Spinosaurus vertebral order and sail shape have been proposed in the last century. While we seem to have a reasonable handle on the arrangement of the anterior sail vertebrae (artists, note that the neural spines project somewhat forward as well as up here: this is a common mistake in spinosaurine art), the shape of the posterior sail slope is more open to interpretation. Originally mounted in the Paläontologische Staatssammlung as a short, tightly-arced sail, Stromer rearranged the vertebrae into a longer, more gently sloping sail in 1936. Later, noting the reclined nature of the posteriormost-known sail spine, others proposed that the sail extended onto the tail (proposed independently by Andrea Cau in 2008 and Jaime Headden in 2010; Paul (2016) shows a similar arrangement while also matching Stromer's 1936 interpretation). More recently, Ibrahim et al. (2014; 2020b) have revived aspects of the pre-1944 Munich arrangement which brings shorter, sometimes anteriorly-positioned spines into a more posterior position (below).
 |
| Various restored shape shapes from a century of Spinosaurus. Images from Smith et al. 2006 and Ibrahim et al. 2014. Be sure to check out other takes on this sail by Andrea Cau, Jaime Headen and Scott Hartman. |
I don't want to pretend that I know which of these arrangements is correct. Arranging these vertebrae is complicated, and there are multiple, perhaps equally viable ways we can order them at present. Based on the new tail data, I suspect the interpretation of Stomer and Ibrahim et al. are correct in restoring the sail plunging sharply into the tail base, but I also see merit to Stomer's 1936 model where vertebra 'f' - the cause of the dip in the Ibrahim et al. model - is positioned more anteriorly.
Unfortunately, the neotype material seems to complicate the resolution of the sail shape further. The sail spines of the neotype are noticeably more slender than those of the holotype despite coming from animals of generally similar size (the neotype is an estimated 11 m long, vs 12 m for the holotype; Dal Sasso et al. 2005; Ibrahim et al. 2020b) and, as preserved, they are quite a bit shorter. Does this imply a lower, less robust sail in the neotype individual, or is this something to do with growth, sexual dimorphism, or another form of variation within Spinosaurus? It's here where our taxonomic assumptions start impacting our reconstructions. Ibrahim et al. (2020a, b) regard S. aegyptiacus as an anatomically variable species, suggesting that we might be OK to blend data from the holotype and neotype sails. Conversely, other schemes regard S. aegyptiacus as potentially confined to Egypt and cast the 'neotype' as a closely related animal (e.g. Evers et al. 2016; Maganuco and Dal Sasso 2018), in which case we might focus more on the sail shape specifically indicated by the Kem Kem specimen. I don't know that there's a clear answer to this conundrum, so artists probably have several options for Spinosaurus sail shapes at present. My own reconstruction follows a somewhat more Stromer 1936-compliant model, as well as a sail height conservatively modelled on the neotype specimen.
Tail flexion
Among the more interesting aspects of the new Spinosaurus tail is the reduction of zygapophyses in the distal region. This potentially allowed the tail to flex far more than was typical for a theropod and to be used for swimming (Ibrahim et al. 2020b). I was initially sceptical of this claim because the long neural spines of the tail extend not only upwards, but also backwards over several other vertebrae, meaning that any movement between vertebrae required the spines to bend in multiple places or else project at wide angles from the tail curve. This is not a novel observation on neural spine length in potentially aquatic animals: I'm basically rehashing arguments made by Silvio Renesto et al. (2010) about the unusual tail of the drepanosaur Hypuronector, and how its extremely long, backwards-projecting chevrons stiffened the tail against sculling-like swimming motions. What I forgot, however, was that Hypuronector also had very developed 'clamping' zygapophyses (Renesto et al. 2010), and what I didn't realise is that - according to folks who know a lot more about biomechanics than I do - the 15 mm wide neural spines of the Spinosaurus tailcould probably bend quite far. The bones of healthy living animals are somewhat plastic and capable of flexion, but I was surprised to learn that muscles and ligaments binding the Spinosaurus tail together would let relatively thick bony rods bend considerably without failing. So perhaps there's less of a problem here than I anticipated, though I admit to wondering how this would work given that Ibrahim et al. (2020b) only reconstruct a very slight covering of soft-tissues on the distal neural spines (below). If muscles only extended up the basal portion of the spine, was this enough to hold the tail together as it sculled the animal through water?
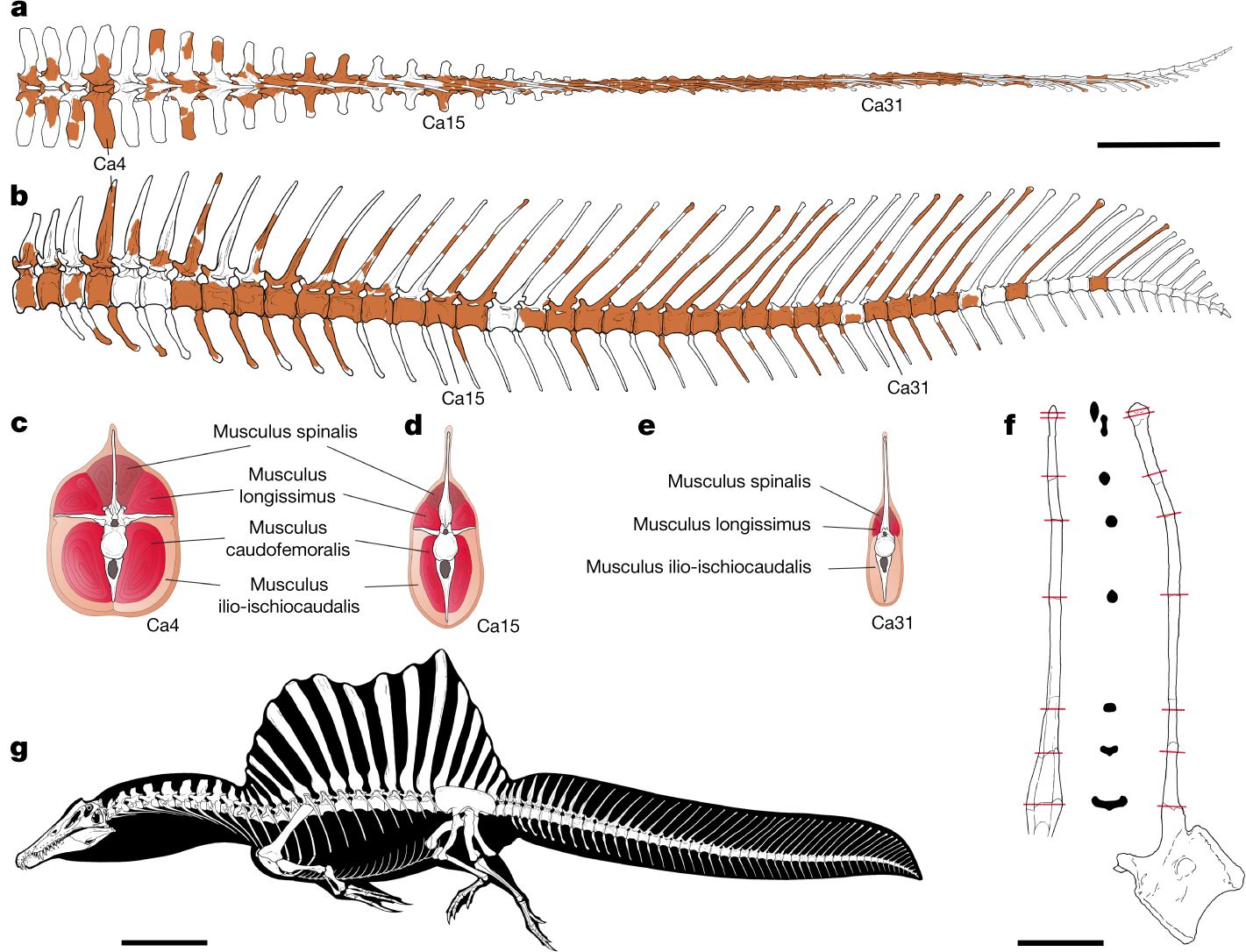 |
| The new tail of Spinosaurus, as presented by Ibrahim et al. (2020b). Note the reduction of musculature in the distal tail ('e') in relation to the discussion of bone bending, above. |
Based in part on these discussions, I've been wondering how much flexibility we can safely reconstruct in the tail. Some of the recent PR imagery has shown a degree of tail flexion that seems beyond that of crocodylians, which seems excessive even allowing for some plasticity in the tail bones. Crocodylian tails have relatively short neural spines and chevrons, as well as large transverse processes to anchor large, strong musculature along much of the tail length. This allows them to pull their tails into tight arcs but, as noted by Ibrahim et al. (2020b), the transverse processes in Spinosaurus are restricted to the anterior tail region in a pretty typical theropod fashion. This musculoskeletal arrangement is thus not very crocodylian-like, and I wonder if the tail was more flexible than usual for a theropod, but maybe not to the degree where it could form a tight, crocodylian-style arc. I also wonder if the energy stored in bending neural spines would spring the tail straight once muscular effort was relaxed, which might have been especially significant when the tail was unrestrained during walking or floating. Maybe, for all its potential flexibility, the tail was held largely straight unless it was actively being used in swimming, or braced against something in the environment.
Facial anatomy and lips
To close out this post, I want to briefly touch on a topic not directly covered in the recent Spinosaurus work, but that comes up whenever spinosaurid illustrations are discussed: did these animals have lipless, crocodylian-like faces? In my experience, lipless spinosaurids are justified by several lines of evidence: their superficially crocodylian-like jaws and teeth; the size and configuration of their anterior teeth (where large premaxillary teeth overbite the lower jaw and long dentary teeth - unusually for a theropod - protrude over the upper jaw during occlusion; Dal Sasso et al. 2005), and the development of liplessness in other semi-aquatic fishers, such as crocodylians and river dolphins.But when looking at spinosaurid jaws with the same criteria generally used to predict extra-oral tissues in fossil animals (tooth size, tooth orientation, jaw bone foramina counts), spinosaurids do not seem unusual compared to other theropods. Their jaws appear peculiar in some ways - check out that foramina rich anterior rostrum, below - because of their atypical geometry, but beyond this, much of their jaw configuration is typically theropodan. Their jaw foramina counts, for example, are not significantly high. Foramina frequency in tetrapod jawbones (premaxilla, maxilla, dentary) have been provisionally hypothesised as indicating the presence of extra-oral soft-tissues in tetrapods (Morhardt 2009), so we can compare foramina counts of Spinosaurus to other tetrapods to infer their facial configuration. Ibrahim et al. (2014) give a Spinosaurus upper jaw foramina frequency of 125, which seems high, but this value represents four bones worth of foramina. Crocodylians have this many foramina, and perhaps many hundreds more, in a single jaw bone. Morhardt (2009) suggests that we need about 100 foramina per jaw bone to infer a lipless condition, which Spinosaurus is well short of. This point recalls comments that the foramina counts and inferred sensitivity of Spinosaurus jaws, which have been correlated to aquatic lifestyles by some authors (Ibrahim et al. 2014), may have been pretty standard for large theropods (Barker et al. 2017), and are possibly not related to aquatic lifestyles or unusual facial anatomy.
 |
| Spinosaurids are often suggested to be among the more likely dinosaurian candidates for liplessness and exposed teeth, but the key features we might look for regarding this condition - labial foramina counts and distribution, as well as jaw bone texture - are not atypical for theropods, nor are they especially crocodylian-like. Their large teeth, including those at the jaw anterior, are no larger (relatively speaking) than those of extant animals with immobile lips and sheathed dentitions (bottom row). Spinosaurus elements after Dal Sasso et al. (2005), Neovenator after Barker et al. (2017); American alligator cropped from original on Wikimedia by Didier Descouens, CC BY-SA 4.0. |
We can also observe that the maxillary and dentary jaw foramina of Spinosaurus are arranged in a more lizard-like row along their oral margins, and not - as in crocodylians - distributed in a dense pattern across the entire jaw. In Spinosaurus at least, they seem to be placed some distance from the toothrow (Dal Sasso et al. 2005) in a lizard-like configuration. This would keep the nerves and blood vessels running into any lip tissues well clear of the overlapping dentary teeth when the mouth was closed, but - based on lizards with similarly displaced foramina - I don't think this means anything too radical for life appearance (lips, if present, would not look unusually big or weirdly anchored). The absence of unusual, epidermally-derived textures on Spinosaurus jaw bones is a further distinction from crocodylians. As we've discussed at lengthin other posts, the characteristic rugosity of crocodylian skulls is reflective of their facial skin and sensory tissues, so the absence of comparable characteristics in Spinosaurus is strong evidence of a different anatomical regime. I'm also not convinced that the teeth of Spinosaurus - so far as they are known (to my knowledge, Spinosaurus jaws with a complete set of teeth remain elusive) - are too large for sheathing behind lips. We have reptiles today with large teeth at their jaw tips and they do not protrude from their lips (above): to the contrary, you'd have no idea they were there from their external appearance.
Finally, what about the purported link between liplessness and fishing aquatic lifestyles? I feel that this reflects a focus on lipless semi-aquatic or aquatic tetrapods but ignorance of the great number of secondarily-aquatic fishers that have retained fully-sheathed dentitions. Yes, crocodylians and river dolphins have unsheathed teeth, but many other fishing swimmers - cetaceans, seals, otters, mink, water monitors, numerous snake species and so on - do not. In fact, many have facial tissues little different to their terrestrial relatives. This questions whether lifestyle is a useful predictor for facial anatomy in Spinosaurus. This is surely a problematic line of evidence anyway, given that it remains to be determined exactly what sort of habits were common to Spinosaurus. Was Spinosaurus an underwater pursuit predator (Ibrahim et al. 2014, 2020b; Gimsa et al. 2016), something more akin to a heron (Hone and Holtz 2015, 2019; Henderson 2016), or something in between? As with so much about Spinosaurus, we have a lot of primary questions to answer before we can start thinking about their implications for behaviour and life appearance.
Enjoy monthly insights into palaeoart, fossil animal biology and occasional reviews of palaeo media? Support this blog for $1 a month and get free stuff!
This blog is sponsored through ;Patreon, the site where you can help online content creators make a living. If you enjoy my content, please consider donating $1 a month to help fund my work. $1 might seem like a trivial amount, but if every reader pitched that amount I could work on these articles and their artwork full time. In return, you'll get access to my exclusivePatreoncontent: regular updates on upcoming books, papers, paintings and exhibitions. Plus, you get free stuff - prints, high-quality images for printing, books, competitions - as my way of thanking you for your support. As always, huge thanks to everyone who already sponsors my work!References
- Allain, R., Xaisanavong, T., Richir, P., & Khentavong, B. (2012). The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos. Naturwissenschaften, 99(5), 369-377.
- Arden, T. M., Klein, C. G., Zouhri, S., & Longrich, N. R. (2019). Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurids. Cretaceous Research, 93, 275-284.
- Barker, C. T., Naish, D., Newham, E., Katsamenis, O. L., & Dyke, G. (2017). Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii. Scientific reports, 7(1), 1-8.
- Benson, R. B., Hunt, G., Carrano, M. T., & Campione, N. (2018). Cope's rule and the adaptive landscape of dinosaur body size evolution. Palaeontology, 61(1), 13-48.
- Dal Sasso, C., Maganuco, S., Buffetaut, E., & Mendez, M. A. (2005). New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its size and affinities. Journal of Vertebrate Paleontology, 25(4), 888-896.
- Evers, S. W., Rauhut, O. W., Milner, A. C., McFeeters, B., & Allain, R. (2015). A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco. PeerJ, 3, e1323.
- Gimsa, J., Sleigh, R., & Gimsa, U. (2016). The riddle of Spinosaurus aegyptiacus’ dorsal sail. Geological Magazine, 153(3), 544-547.
- Henderson, D. M. (2018). A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic Spinosaurus Stromer, 1915 (Dinosauria: Theropoda). PeerJ, 6, e5409.
- Hendrickx, C., Mateus, O., & Buffetaut, E. (2016). Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa. PLoS One, 11(1).
- Hone, D. W. E., & Holtz Jr, T. R. (2017). A century of spinosaurs‐a review and revision of the Spinosauridae with comments on their ecology. Acta Geologica Sinica‐English Edition, 91(3), 1120-1132.
- Hone, D. W. E., & Holtz Jr, T. R. (2019). Comment on: Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurids. 93: 275–284. Cretaceous Research, 104152.
- Ibrahim, N., Sereno, P. C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D. M., ... & Iurino, D. A. (2014). Semiaquatic adaptations in a giant predatory dinosaur. Science, 345(6204), 1613-1616.
- Ibrahim, N., Sereno, P. C., Varricchio, D. J., Martill, D. M., Dutheil, D. B., Unwin, D. M., ... & Kaoukaya, A. (2020a). Geology and paleontology of the Upper Cretaceous Kem Kem Group of eastern Morocco. ZooKeys, 928, 1.
- Ibrahim, N., Maganuco, S., Dal Sasso, C., Fabbri, M., Auditore, M., Bindellini, G., ... & Wiemann, J. (2020b). Tail-propelled aquatic locomotion in a theropod dinosaur. Nature, 1-4.
- Lakin, R. J., & Longrich, N. R. (2019). Juvenile spinosaurs (Theropoda: Spinosauridae) from the middle Cretaceous of Morocco and implications for spinosaur ecology. Cretaceous Research, 93, 129-142.
- Maganuco, S., & Dal Sasso, C. (2018). The smallest biggest theropod dinosaur: a tiny pedal ungual of a juvenile Spinosaurus from the Cretaceous of Morocco. PeerJ, 6, e4785.
- Morhardt, A. C. (2009). Dinosaur Smiles: Do the Texture and Morphology of the Premaxilla, Maxilla, and Dentary Bones of Sauropsids Provide Osteological Correlates for Inferring Extra-oral Structures Reliably in Dinosaurs?: A Thesis. Western Illinois University.
- Paul, G. S. (2016). The Princeton field guide to dinosaurs. Princeton University Press.
- Renesto, S., Spielmann, J. A., Lucas, S. G., & Spagnoli, G. T. (2010). The taxonomy and paleobiology of the Late Triassic (Carnian-Norian: Adamanian-Apachean) drepanosaurs (Diapsida: Archosauromorpha: Drepanosauromorpha): Bulletin 46 (Vol. 46). New Mexico Museum of Natural History and Science.
- Smith, J. B., Lamanna, M. C., Mayr, H., & Lacovara, K. J. (2006). New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915. Journal of Paleontology, 80(2), 400-406.
- Taylor, M. P., & Wedel, M. J. (2013). Why sauropods had long necks; and why giraffes have short necks. PeerJ, 1, e36.