![]() |
| Deinosuchus rugosus swallows the remains of a large Cretaceous sea turtle. Other archosaurs notice, decide to interrupt. |
Any palaeontological geek worth their salt knows that several gigantic crocodyliform species - colloquially called 'supercrocs' - have appeared in the last 100 million years. They include the Moroccan, Cretaceous pholidosaurid
Sarcosuchus imperator, several species of the South American Miocene caimanine
Purusaurus, and the grandfather of them all,
Deinosuchus, from the Late Cretaceous (Campanian) of North America.
Deinosuchus is not the most 'extreme' of these giant crocodyliforms in terms of anatomy or size (but see below), but discovery of a partial skeleton in 1903 (and description six years later) was the first evidence for some croc-line archosaurs being very, very large. It is also perhaps the best publicly known giant crocodyliform, various reconstructions of its skull and skeleton appearing in museums all over the world, and being a semi-regular component of palaeoart.
Perhaps because
Deinosuchus is 'only' a giant crocodylian and not a member of a completely extinct, weirdo lineage, coverage of its palaeobiology is often limited to factoids on its immense size and probable habits of eating dinosaurs. Our short attention span for this animal is not new: one of the key players in its discovery, John Bell Hatcher, lost interest in describing the first significant
Deinosuchus remains once its crocodylian identity (rather than dinosaurian, as originally supposed) became apparent in 1903 (Schwimmer 2002). It took further persuasion and a number of years for palaeontologists to actually publish, name and describe this animal after Hatcher died in 1904 (Holland 1909). As we'll discover in this article, the scant attention paid to this animal is rather criminal: over the last century a detailed and fascinating picture of
Deinosuchus has developed.
![]() |
| The Colbert and Bird (1954) reconstruction of Deinosuchus riograndensis, based on partial skull material collected from the late Campanian Aguja Formation, Texas. We now appreciate this reconstruction as being erroneous in a number of crucial ways, and it should not be considered representative of the appearance of this animal. Note the apertures in the snout tip alongside the actual nares. Images from Colbert and Bird (1954). |
I'm going to start by sidestepping the confused history surrounding the discovery and naming of
Deinosuchus material - interested parties can find a full summary in David Schwimmer's (2002) book
Deinosuchus: King of the Crocodylians. It will suffice to state that fossils of this animal have been known since at least the mid-1800s and repeatedly hopped between species and genera during the last 150 years. Only one or two species are recognised nowadays.
D. rugosus is the type species of the genus, first identified from two characteristically large, blunt teeth with wrinkled, thick enamel collected from the eastern US in the 1858 (examples below). Over time, these teeth were found to be linked with other
Deinosuchus material including extremely thick, massive and deeply pitted osteoderms. These elements are highly characteristic even today, and permit even isolated teeth and osteoderms to be referred to this species (Schwimmer 2002). A possible second species is
D. riograndensis, based on very large
Deinosuchus fossils from Texas recovered in the 1940s (Colbert and Bird 1954). It's this
riograndensis material that most of us think of when
Deinosuchus is mentioned, it providing the basis of a famous, 2 m long and largely artistic
Deinosuchus skull reconstruction unveiled at the AMNH in 1954 (above). The
riograndensis skull was once thought characteristic because of unusual openings in the side of the snout tip (Colbert and Bird 1954), but these are now thought to be damage caused during preparation (Schwimmer 2002). As more and better
Deinosuchus remains have been recovered in recent decades, some have argued
riograndensis and
rugosus are one and the same animal, the latter holding nomenclatural priority. Full agreement on this does not seem apparent from current literature, but on-going work on relatively complete
Deinosuchus material will help clarify the taxonomy of this animal in future.
![]() |
| Deinosuchus is recognisable from its teeth alone. These specimens are from the posterior end of the jaws, and show the wrinkled enamel typical of the genus. Note the heavily worn and broken the tips. From Schwimmer (2010). |
We now appreciate that
Deinosuchus is one of the oldest members of Crocodylia, the crown group crocodile-line archosaurs. Features of its skull and jaws indicate it was specifically a member of Alligatoroidea, the lineage of Crocodylia represented today by alligators and caimen. It's not uncommon to see
Deinosuchus referred to as a 'giant alligator', but this is not really accurate and I recommend avoiding such terminology, even for lay audiences. Alligators and Alligatoridae are a distinctive group of alligatoroids with particular habits and anatomy, and they are no more closely related to
Deinosuchus than caimen. It must also be stressed that neither of our modern alligatoroid clades are especially closely related to
Deinosuchus. I figure most people will intuitively grasp the rough meaning of 'giant alligatoroid', and suggest this term is used in preference of 'alligator' in outreach media about this animal.
The
Deinosuchus fossil record is something of a mixed bag. There are hundreds of fossils of it, but most of them are isolated postcranial bones, broken bits of skull and, especially, those massive teeth and osteoderms. The state of many
Deinosuchus fossils can be ascribed to its remains being reworked by storms after their initial burial. Some partial skeletons and more complete skulls escaped this treatment but are not yet described in detail. A silver lining to not having much in the way of complete material is that isolated
Deinosuchus bones are distinctive enough to map its range across Campanian North America. Many of us might think of
Deinosuchus as a Texan animal, but it actually enjoyed a wide distribution, and being most abundant in the southeastern United States. A clear palaeobiogeographical pattern can be gleaned from
Deinosuchus fossils, a divide separating occurrences in Montana, Wyoming, Utah, Colorado, New Mexico, Texas, and Northern Mexico from remains on the eastern side of the United States - Mississippi, Alabama, Georgia, North Carolina, and New Jersey (Titus et al. 2008). This east-west distribution is no fluke of preservation but reflection of
Deinosuchus populations being separated by the Western Interior Seaway, the continental sea which divided North America during the Cretaceous. Although apparently not a fully marine creature, it is thought
Deinosuchus lived in the coastal waters and estuaries of this seaway as, to date, its fossils have not occurred in fully freshwater or terrestrial deposits. Further evidence of its preference for coastal waters is the recovery of more complete and associated remains from wholly marine deposits.
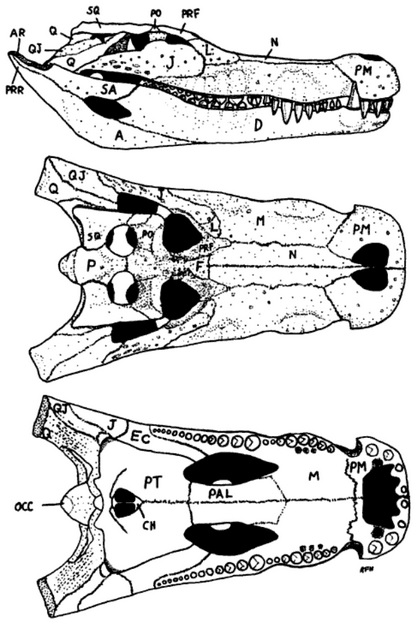
![]() |
| A modern depiction of the Deinosuchus rugosus skull and mandible. From Schwimmer (2002). |
Our incomplete knowledge of
Deinosuchus anatomy means we can only form a partial picture of what it looked like in life. We can say, however, that the famous 'Colbert and Bird'
riograndensis skull sculpture from the 1950s is erroneous in several regards. Manufactured as a display for the American Museum of Natural History (and then reproduced for museums around the world), the Colbert and Bird skull is too large, the snout too narrow, the tooth morphology inexact and, as noted above, the openings in the snout tip are likely erroneous (Schwimmer 2002). This is not a dig at AMNH artists course: they did the best they had with material available to them, and it's only with the discovery of better fossils that we can now spot errors. Perhaps unfortunately, museums continue to display this reconstruction and artists continue to use it as a reference. A modern picture of
Deinosuchus is rather different: a broad- and deep-snouted crocodylian with a skull known to be at least 1.3 m in length, and perhaps a little longer if very fragmentary remains are being correctly interpreted (Schwimmer 2002). Most of its cranial features are typically crocodylian, including the presence of huge spaces for jaw muscle attachment, development of a secondary palate and dorsally situated orbits and nares. Befittingly for such an enormous animal, the teeth are huge and consistently robust along the jaw, those at the front being conical and pointed, and those at the back being increasingly stunted and shortened. The toothrow is very long in stretching from the jaw tip to just behind the eye. All teeth have the distinctively thickened, wrinkled enamel mentioned above. Unlike modern alligatoroids, a notch in the side of the upper jaw acts as a receptacle for the fourth tooth of the lower jaw and presumably rendering it visible even when the jaw was closed. This is the 'primitive' condition for crocodylians, and means that although
Deinosuchus is an alligatoroid, it likely had a crocodile's smile.
Less can be said about the body and limbs of
Deinosuchus. Schwimmer (2002) reports that partial skeletons hint at a general form and proportion not unlike a modern alligator. However, some authors have noted discrepancies in scaling of
Deinosuchus limb bones which might indicate reduced limbs in at least the largest specimens (Farlow et al. 2005, see below). One thing we can be sure of is that the body of
Deinosuchus was covered in those aforementioned large, thickened osteoderms (below). The exact arrangement of these elements remains unknown, but we can predict that at least four rows of osteoderms extended along the body of
Deinosuchus because of its affinities to modern crocodylians. These osteoderms become disproportionately massive and robust with growth, so that those of the largest individuals are distinctively chunky and have lost some definition of a keel found in smaller examples. Artists should take note of this: the dorsum of a big
Deinosuchus would have looked more like a gnarly
Dalek chassis than the back of any modern crocodylian. As is typical for crocodyliforms, these dermal bones might have reinforced the trunk skeleton as well as providing armour plating, forming a network of muscle, ligaments and bone which bound the torso together (Salisbury and Frey 2000). It is speculated that the presence of very large, robust osteoderms in the biggest
Deinosuchus indicates the presence of a torso strong enough for terrestrial locomotion (Schwimmer 2002).
![]()
|
| The huge, deeply pitted and bulbous scutes which characterise Deinosuchus, as illustrated by Holland (1909). This image is a composite of two scutes from Holland's work, put together by FanCollector for Wikipedia. Both show cervical osteoderms, the left being a particularly big one |
The maximum size of
Deinosuchus is the source of much fascination and discussion. The largest estimates based on reliably measured remains suggest body lengths of around 12 m (Schwimmer 2002), but - as usual with giant fossil animals - there are a number of factors and caveats worth considering here. The scant nature of
Deinosuchus fossils dictates that we must extrapolate the size of large
Deinosuchus from much smaller, better known individuals and modern crocodylians. The 12 m figure stems from scaling the largest
Deinosuchus vertebrae to total body length estimates of smaller individuals (Schwimmer 2002). Skulls and mandible lengths, when compared to a dataset of modern alligator proportions, indicate the largest animals achieved 10 m in length (Schwimmer 2002; Farlow et al. 2005). Another approach, using femoral measurements, results in a maximum body length estimate of 6-8 m, this being estimated from big femora typically thought to indicate 10 m+ animals. The explanation for these differences will be familiar to anyone who's estimated the size of a big, extinct animal: uncertainty about the proportions of the animal in question, the need to extrapolate well beyond the size boundaries of modern analogues, and the lack of associated remains of the biggest individuals. Most workers seem happy with a 10 m length estimate for a big
Deinosuchus, those lower estimates based on femoral size being explained as possible evidence of reduced limb proportions in the biggest
Deinosuchus individuals (Farlow et al. 2005). As usual, we await the discovery of more complete and informative remains to tell us the full story here. It should be stressed that similar caveats apply to size estimates of other 'supercrocs': despite the media hype associated with some discoveries, it's quite difficult to know which crocodyliform species was largest based on our current material.
Regardless of what the actual maximum size of
Deinosuchus was, we have good reason to think that many individuals were not true giants. No specimens indicative of 10 m body length have been found among the many hundreds of
Deinosuchus remains from the eastern side of the US, it being instead thought that eastern
Deinosuchus didn't grow longer than 8 m. That's still pretty big of course, but not too far off crocodylians that we're familiar with today (the biggest saltwater and Orinoco crocodiles on record are a little over 6.5 m - Grigg and Krishner 2015). The true giants only occur in the west, and are much rarer fossils than their eastern counterparts. These fossils are also slightly younger than the eastern specimens, perhaps indicating changes across time and geography were responsible for
Deinosuchus becoming exceptionally large. Research into the growth rates of
Deinosuchus indicate that there might be nothing unusual about it's growth trajectory despite its size. It seems to have grown with a similar strategy to other crocodylians - relatively fast at first, and progressively slower over time - but simply stretched out the growth duration to many decades (Erickson and Brochu 1999). Growth rings in osteoderms indicate that the largest animals were about around 50 years old (below).
![]() |
| Growth rates in living and extinct crocodylians (a) and growth rings in a Deinosuchus osteoderm (b). Note how the Deinosuchus growth trajectory is essentially a scaled up version of its smaller relatives. From Erickson and Brochu (1999). |
For many, the main discussion to be had about
Deinosuchus is the impact it had on local dinosaur populations: was this animal a dinosaur predator? Attempts to answer this question stem from two sources: biomechanics and fossil evidence of ancient faunal interactions. The first, biomechanics, includes a recent study of 'death rolling' (the crocodylian habit of rotating around the long axis of the body while gripping prey with their jaws, literally twisting it apart) and whether
Deinosuchus could use this strategy to dismember large prey (Blanco et al. 2015). Snout strength can be correlated quite accurately to death rolling capabilities in modern crocodylians (Blanco et al. 2015) and, seeing as this can be inferred from upper jaw skeletons alone, we can obtain some insight into the death rolling capabilities of extinct crocodyliforms. Perhaps surprisingly, estimates of
Deinosuchus jaw strength were approximately one third of the strength required for this behaviour (Blanco et al. 2015). Scaling factors have been used to explain this unexpected result and, despite the outcome of their experiments, Blanco et al. suggest that death rolling was possible in
Deinosuchus. I must admit to thinking additional experimentation is needed to quantify those scaling factors before considering this matter closed. Moreover, in checking the Blanco et al. data for this article I noted that their study modelled the
Deinosuchus skull as 1.8 m long, a figure noted as speculative by Schwimmer (2002) and almost 40% longer than the largest measured skull length reported by the same author (1.31 m). A 40% shorter skull would be less prone to the scaling effects outlined by Blanco et al. and may result in a jaw strength more suited to death rolling - it would be great to see this checked out in future.
![]() |
| Partial theropod hindlimb bone (tibia or metatarsal) post a one-on-one session with Deinosuchus jaws. This bone is meant to be subrounded in cross section. From Schwimmer (2010). |
More positive and definitive answer about
Deinosuchus dinosaur predation stems from fossil evidence. It seems that, yes,
Deinosuchus did dine on dinosaurs, but not exclusively or maybe even often (Schwimmer 2010). A handful of Campanian dinosaur bones - including a theropod hindlimb element from Georgia (above) and hadrosaur vertebrae from Texas - possess bitemarks characteristic of crocodylians, albeit on a scale unseen in the modern day (Schwimmer 2002, 2010). The theropod bone can only be described as exceptionally chewed: numerous, overlapping circular potmarks show where the bone was repeatedly bitten and crushed by a very powerfully jawed animal. So pulverized is this bone that its once subrounded cross section has become quadrangular - Schwimmer (2002) summarises the state of this specimen as 'resembling a dog's worn chew toy' (p. 186). The crocodylian signature, bite mark size and provenance of both specimens point to
Deinosuchus as a possible perpetrator, and evidence that it did eat dinosaurs on occasion. Of course, it cannot be easily established whether these animals were killed by
Deinosuchus or merely scavenged by them - some reasoning for the former is discussed by Schwimmer (2010).
![]() |
| Deinosuchus bite marks in fragments of a turtle (Chedighaii barberi) plastron. The tooth marks are about 4-5 times larger than those made by 4 m long nile crocodiles. From Schwimmer (2010). |
|
|
There are reasons to think dinosaur meat was not a mainstay of
Deinosuchus diet, however. Whereas a few dinosaur bones have been linked to the jaws of this crocodylian, more than a dozen Campanian sea turtle specimens have been found with bite marks made by a giant crocodylian (Schwimmer 2010). A diet of turtles is not surprising when we consider that the skull and dentition of
Deinosuchus is more adapted to crushing bone than piercing skin and flesh (Schwimmer 2002): those robust posterior teeth are especially reminiscent of teeth in modern, turtle-eating crocodylians. Many
Deinosuchus teeth are considerably worn and broken too, a likely consequence of being smashed into hard, bony prey rather than soft, spongy dinosaur limbs. Healed turtle shells suggest that these animals were predated by
Deinosuchus rather than just scavenged, and raking bites across some specimens may record less fortunate turtles being juggled about
Deinosuchus jaws when being eaten (Schwimmer 2010). The coastal and estuarine environmental bias of
Deinosuchus fossils is consistent with it being a serial turtle predator, this being ideal habitat to find sea-going prey. Curiously, other marine inhabitants of the Western Interior Seaway have yet to be associated with
Deinosuchus bite marks: perhaps it really did have a preference for turtles, or perhaps other skeletons were simply pulverised beyond recognition by those massive jaws. Either way, our discussion of this animal's feeding habits would not be complete without mentioning the numerous, possible
Deinosuchus coprolites which have recently been identified (Harrell and Schwimmer 2010). Sadly, these do not reveal much about diet or digestive anatomy, other than the obvious fact that
Deinosuchus poop was on average a lot larger than that produced by other crocs. Several anomalous features of these coprolites have led some authors (e.g. Hunt and Lucas 2010) to be sceptical of their organic origins however, their alternative being that they are simply calcareous nodules.
The foraging habits of
Deinosuchus brings us to new perspectives on where it fits into Mesozoic ecology. New evidence is eroding the uniqueness of
Deinosuchus in Campanian North America, it no longer being the only very large or even giant crocodyliform species in some localities. These new finds include a currently poorly known, but obviously giant neosuchian from the Williams Fork Formation of Colorado (Foster and Hunt-Foster 2015) and a more completely known, 7 m long, undescribed neosuchian from Woodbine, Texas (Main 2012). The latter is currently being worked on, and early indications are that it might represent a late surviving
goniopholidid - a much older branch of the crocodyliform lineage. Whatever they turn out to be, the Woodbine and Williams Fork animals suggest that very large crocodyliforms might not have been unusual in Campanian North America. Their presence in a timeframe deficient of large theropods has not gone unnoticed, it being speculated that these large crocodyliforms may have been doing work normally reserved for big predatory dinosaurs (e.g. Schwimmer 2002). Similar proposals have been made about other large bodied Late Cretaceous carnivores taking over typically theropodan roles (e.g. Witton and Naish 2015) - the notion of the Mesozoic as an all-dinosaur show is looking increasingly out of date.
As a closing thought, I find it interesting that we tend to portray
Deinosuchus as something of a freak species, one of those rare forays of crocodylian evolution into gigantic size which never really seemed to last that long or lead anywhere. As might be apparent from this article, this view is somewhat misleading.
Deinosuchus certainly represents an 'extreme' of crocodylian evolution, but it's at the end of a spectrum, not a weird outlier from the rest of the group. Much of what it did, how it did it, and what makes it a fascinating animal, is mirrored in its modern and fossil relatives. Contrary to some perspectives on this animal, the fact it represents an ancient member of a modern group does not make it tedious or dull. Quite the opposite is true:
Deinosuchus reminds us that animals from Deep Time are part of a continuum with our own fauna, revealing the awesome things modern lineages have been capable of, the potential their anatomies have in the present, and what they might be up to in future. How anyone can find pondering an animal that gives such a raw perspective on evolution and adaptation boring or uninteresting is beyond me.
Now is a great time to sign up to my Patreon
The artwork and words you see here are supported by Patreon, the service which allows you to directly support artists and authors with monthly payments. You can support this blog from $1 a month and, in exchange, get access to bonus content and rewards - the more you pledge, you more bonuses you receive.
In February be sending out signed art prints to all those who pledge at least $5 a month - sign up now to get yourself on the receivership list!
References
Blanco, R. E., Jones, W. W., & Villamil, J. (2015). The ‘death roll’of giant fossil crocodyliforms (Crocodylomorpha: Neosuchia): allometric and skull strength analysis. Historical Biology, 27(5), 514-524.
- Colbert, E. H., Bird, R. T., & Brown, B. (1954). A gigantic crocodile from the Upper Cretaceous beds of Texas. American Museum Novitates; no. 1688.
- Erickson, G. M., & Brochu, C. A. (1999). How the ‘terror crocodile’ grew so big. Nature, 398, 205-206.
- Farlow, J. O., Hurlburt, G. R., Elsey, R. M., Britton, A. R., & Langston Jr, W. (2005). Femoral dimensions and body size of Alligator mississippiensis: estimating the size of extinct mesoeucrocodylians. Journal of Vertebrate Paleontology, 25(2), 354-369.
- Foster, J. R., & Hunt-Foster, R. K. (2015). First report of a giant neosuchian (Crocodyliformes) in the Williams Fork Formation (Upper Cretaceous: Campanian) of Colorado. Cretaceous Research, 55, 66-73.
- Grigg, G., & Kirshner, D. (2015). Biology and Evolution of Crocodylians. Csiro Publishing.
- Harrell, S. D., & Schwimmer, D. R. (2010). Coprolites of Deinosuchus and other crocodylians from the Upper Cretaceous of western Georgia, USA. New Mexico Museum of Natural History and Science, Bulletin, 51, 209-213.
- Holland, W. J. (1909) Deinosuchus hatcheri, a new genus and species of crocodile from the Judith River beds of Montana. Annals of the Carnegie Museum, 6, 281–294.
- Main, D. J. (2012). Crocodiles of the Texas Cretaceous; the Campanian of Big Bend to the Cenomanian of North Texas, a comparison of great size, feeding behaviour and paleoecology. Geological Society of America Abstracts with Programs, 44, 3.
- Schwimmer, D. R. (2002). King of the crocodylians: the paleobiology of Deinosuchus. Indiana University Press.
- Schwimmer, D. R. (2010). Bite marks of the giant crocodylian Deinosuchus on Late Cretaceous (Campanian) bones. New Mexico Museum of Natural History and Science Bulletin, 51, 183-190.
- Titus, A.L., Knell, M.J., Wiersma, J.P., Getty, M.A. (2008). First report of the hyper-giant Cretaceous crocodylian Deinosuchus from Utah. Geological Society of America Abstracts with Programs, 40, 58.
- Witton, M. P. and Naish, D. (2015) Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"? Acta Palaeontologica Polonica 60, 651-660.