![]() |
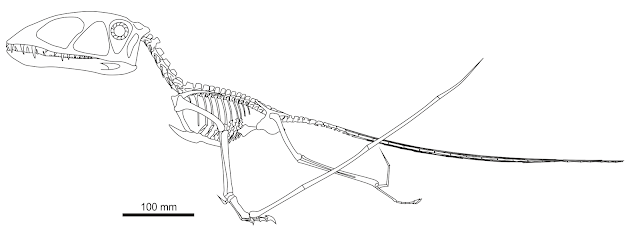
| The new Tanystropheus cf. longobardicus skeletal reconstruction I presented in my last post. What the dickens did this crazy animal do? That's what we're discussing today. |
What sort of animal was the Triassic, long-necked Eurasian protorosaur
Tanystropheus? As we discovered in the last post, the lifestyle of
Tanystropheus remains controversial over a century after it was first discovered. There is near universal agreement that it ate swimming prey such as fish and squid, but opinion is divided over whether it was obligated to aquatic, swimming lifestyles because of the burden of its long neck, or whether it was a water margin specialist that plundered small prey from shorelines.
Previously, we discussed a core argument for the aquatic hypothesis, that the
Tanystropheus neck would over-balance the animal. Calculations presented in the last post suggested that the mass distribution of
Tanystropheus is not as weird as we might think, and certainly less so than than that of another group of long necked reptiles we are confident lived out of water, the
azhdarchid pterosaurs. Based on this very basic test, I expressed some skepticism about the neck being simply too heavy to permit a terrestrial existence.
In the second discussion, I want to look at some finer aspects of
Tanystropheus anatomy and palaeontology, how they've been interpreted, and what they might mean for its lifestyle. There are several areas which are relevant here: what we know of
Tanystropheus diet, the palaeoenvironmental context of
Tanystropheus fossils, aspects of tail and limb anatomy, and of course, the functionality of its neck. There's a lot to get through here, so let's not waste any more time on preamble.
Fossil record
An obvious line of inquiry about ancient animal habits is the palaeoenvironmental bias of its fossil remains, and the fossil organisms it is found with. We mentioned last time that
Tanystropheus was a wide-ranging taxon, occurring across Europe, Israel and China in locations representing the coasts and shallow waters around the ancient Tethys ocean. About half of
Tanystropheus fossils come from shallow marine settings, the rest being derived from more coastal environments: river and estuarine environments, lagoons, intertidal settings and so forth (for a brief overview, check out the
Fossilworks entry on this animal: there's a few localities missing, and the 'terrestrial' occurrence of
Tanystropheus there is erroneous, but it gives a flavour of its depositional context). We often find marine fish and seagoing reptiles in the same beds as
Tanystropheus,
but it also occurs alongside terrestrial or freshwater species such as temnospondyls, terrestrial reptiles, stem mammals and plants in a number of locations. The link of
Tanystropheus to these faunas seems complex: in at least one locality with fluctuating marine and terrigenous influences,
Tanystropheus fossils only occur in horizons containing a mix of highly terrestrial and highly marine reptiles, without many 'intermediate' semi-aquatic species (Renesto 2005). Because
Tanystropheus was likely not adapted for a truly seagoing lifestyle, this has been argued as evidence of it being part of a terrestrial community rather than a marine one (Renesto 2005).
Collectively, it seems difficult to argue a strong terrestrial or marine bias in this record.
Tanystropheus seems to have lived in or around aquatic environments, maybe with a bias to those under marine influences, but it does not seem a stranger to brackish or freshwater settings. There is perhaps something of a skewed association with marine animals, but it occurs with enough 'terrestrial' forms to keep the idea of a coastal fishing lifestyle buoyant. It would be interesting to put some actual numbers on this and see how commonly associated with terrestrial influences
Tanystropheus is, or whether a couple of sites are skewing our perception of data. Maybe that's a job for another blog post - until then, we probably need to look at other sources of information for clearer lifestyle indications.
Gut content
The idea that Tanystropheus ate swimming prey is verified by the association of digested fish remains and cephalopod hooks in the gut regions of articulated specimens (Wild 1973; Li 2007). The latter is sometimes considered smoking gun evidence for the swimming
Tanystropheus lifestyle hypothesis, it being reasoned that cephalopods are exclusively marine animals, mostly found far out to sea, and unlikely to be eaten from land (e.g. Nosotti 2007).
![]() |
| A number of heron species, including the globally distributed black-crowned night heron (Nycticorax nycticorax), are known squid-eaters. Image from Wikimedia (CC ), by Kuribo. |
Squiddy gut content certainly matches the idea of a marine-influenced lifestyle for Tanystropheus, but several non-marine, and sometimes non-aquatic, birds and mammals challenge the idea that it had to be a swimming animal to have ingested them. Examples include night herons (Hall and Cress 2008) and several types of mustelid (e.g. Hartwick 1983; Beja 1991). Exactly how night herons obtain squid is not documented in detail, but photographs of twoother heron species demonstrate squid can be apprehended without venturing out to sea, or even into deep water. As might be expected, cephalopods also frequently wash up on beaches (sometimes still alive, and in huge numbers) allowing animals such as bears and wolves to also access cephalopod meat. Humans are also adept predators of squid in coastal settings. Shore-based squid angling is reportedly a growing hobby around the world (and apparently requires only very basic fishing equipment) and we routinely collect cephalopods from intertidal environments for use as bait or cooking ingredients (Denny and Gains 2007). Contrary to expectations, accessing cephalopod prey from shore environments appears quite possible for a number of differently adapted species. It seems premature to rule out a coastal fishing lifestyle for Tanystropheus just because it sometimes ate squid-like animals. Anatomy
![]() |
| One of the most famous and complete Tanystropheus longobardicus specimens known, MSNM BES SC 1018. This illustration is from Nosotti's huge (2007) monograph. |
With the fossil record and gut content providing slightly ambiguous insight into
Tanystropheus habits, its functional anatomy is probably going to be a deciding card here. A lot has been said about the functional morphology of
Tanystropheus, and there is a lack of consensus on many issues. For instance, its neck flexion has been described as almost 'swan-like' (Wild 1973); broom handle-stiff (Tschanz 1988), or somewhere inbetween (Renesto 2005). Its tail has been considered lousy for aquatic propulsion by some (Wild 1973; Renesto 2005) but well suited for the job by others (Tschanz 1988; Nosotti 2007). Clearly, some of these ideas must be erroneous, them being too polarised for all contributing parties to be correct. Such confused functional interpretations are not without precedent: Darren Naish and I noted a similar situation with azhdarchid pterosaurs in our 2008 paper: maybe this is simply what happens when we try to understand weird fossil species.
The main points of contention about
Tanystropheus functional anatomy concern its tail, limbs and neck. We might link these attributes to two principle functions: locomotion and foraging. Let's start with the former. Proponents of the aquatic
Tanystropheus hypothesis suggest the tail was the likely propulsive organ, it being considered that the limbs are too long and gracile to function as effective paddles (Tschanz 1988; Nosotti 2007), even if the foot might have some aquatic adaptations (below; Kuhn-Schnyder 1959; Wild 1973). Near 'horizontal' articulations between the posterior trunk and tail vertebrae appear to have permitted this part of the body to undulate laterally, permitting a crocodile-like sculling approach to swimming.
![]() |
| Soft-tissue preservation around the tail of Tanystropheus cf. longobardicus specimen MCSN 4451. We're looking at the underside of the tail in the left of the image here - note the width of the soft-tissue (the big grey mass). The verts on the right are shown in left lateral view. From Renesto (2005). |
A fly in the ointment here is the gross tail anatomy of
Tanystropheus. Rather than being long, and comprised of the robust, tall vertebrae expected of a tail-propelled aquatic reptile, its tail is slender, relatively short and actually broader than tall - hardly an ideal sculling organ (Renesto 2005). This fact has been noted by proponents of the swimming lifestyle hypothesis, and it has been proposed that the tail sported some sort of fin to modify it into a swimming organ (Nosotti 2007). Well, maybe, but this idea is entirely without support from fossil data. Readers may recall that marine reptile workers have been quite ingenious in their ability to detect fins and flukes from osteological correlates, none of which are obvious in the tail of
Tanystropheus. Moreover, preserved soft-tissues from the anterior
Tanystropheus tail region (above) show no signs of fins but instead a broad tail base unconducive to aquatic propulsion (Renesto 2005). Also worth mentioning is recent work on the relationship between vertebral articulation and swimming capability in crocodyliforms. They
can reflect sculling behaviour, but articulations like those seen in
Tanystropheus can also be linked to preventing trunk collapse during non-aquatic locomotion (Molnar et al. 2014). We could go on, but I think the point has been made that arguments for the
Tanystropheus tail being a swimming organ are, at best, not without complication, and perhaps better described as uncompelling.
Turning our attention to the limbs, I mentioned in the last post that I was surprised how 'leggy'
Tanystropheus was when restored as walking rather than,
as we're used to seeing it, squatting. The limb proportions and girdle sizes of
Tanystropheus compare well with non-aquatic protorosaurs such as
Macrocnemus and
Langobardisaurus (e.g. Renesto 2005; Nosotti 2007) and, as alluded to above, it is immediately clear that these limbs are not flippers. Not only are they too long and gracile for effective use as hydrofoils, but their long bones are hollow - unexpected features of an aquatic animal. Another protorosaur -
Dinocephalosaurus - gives an insight into how these reptiles could modify their limbs into efficient flippers (below), and, without going into detail, they're nothing like the limbs of
Tanystropheus (see Renesto 2005 for a long discussion of this).
Tanystropheus limb joints are mostly robust and well-defined (but see below), and its hands and feet are strongly built and compactly structured. Some differences between hand and foot proportions can be seen: the hands are short, the feet rather long, and the latter characterised by a peculiarly long first bone in the fifth toe. The limb girdles are well developed, looking proportionally comparable (speaking from pure eyeballing here, not precise measurements) to those of large monitor lizards and crocs. I find the shoulder blade of particular interest, as it is rather large and broad, subequal in proportions to the coracoid (the lower portion of the shoulder girdle). This contrasts with many aquatic animals, which tend to maximise the size of the coracoids while reducing the scapula.
![]() |
| Variations in protorosaur limb anatomy, demonstrated by the aquatic Dinocepahlosaurus (A-B) and Tanystropheus (C-D). Note how both the arm (A) and leg (B) of Dinocephalosaurus are short and wide compared to their equivalents in Tanystropheus (forelimb = C, hindlimb = D), making them much more effective flippers. You can also see the reduced mineralisation in the Tanystropheus wrist here. From Renesto (2005). |
I have to agree that
Tanystropheus limbs were probably unchallenged by non-aquatic habits (Renesto 2005) and, if this were any other species, I don't think we'd be disputing the fact that its limbs were likely capable of terrestrial locomotion. That said, there are undeniably some hints that
Tanystropheus was not
always walking on land. Several authors have noted that the wrist and ankle bones of
Tanystropheus are not as well ossified as those of other protorosaurs (e.g. Rieppel 1989; Nosotti 2007), and some have suggested that the pelvic bones may also be somewhat less defined (Rieppel 1989). Moreover, the elongation of the fifth toe is atypical for a purely terrestrial reptile, but common among aquatic creatures (see Kuhn-Schnyder 1959 for a good illustration of this point). Proposals that this made the foot somewhat paddle-like, or supported
Tanystropheus on soft, saturated substrates do not seem unreasonable. These are fairly minor modifications to the skeleton when viewed overall however: the reduced ossification in the wrist, ankle and pelvis is pretty minor - especially when we consider how cartilage-filled the joints of many giant terrestrial archosauromorphs can be (Holliday et al. 2010) - and the reconfiguration of foot bones do not override the otherwise elongate, gracile structure of the hindlimb. My overall interpretation of the limb configuration broadly agrees with that proposed by Renesto (2005): a bauplan suited to terrestrial locomotion with some aquatic leanings, rather than sustained aquatic propulsion.
Finally, we come to the neck. I've saved discussion of this for last because I consider much of its anatomy significant in terms of where
Tanystropheus lived and how it accessed food. Discussing it earlier might have rendered other points a bit superfluous. We make a lot of noise about how strange the neck of this animal is, but
Tanystropheus neck anatomy frequently converges with those of other long necked reptiles - pterosaurs and sauropods - and even some long-necked mammals. That doesn't necessarily make it less weird - it's definitely still an 'extreme' biological structure - but does help us put its neck anatomy in perspective with other animals, as well as highlighting significant adaptive differences to neck elongation in aquatic and non-aquatic species.
As with pterosaurs and sauropods,
Tanystropheus went to great lengths to lighten its neck. Firstly, its neck is comprised of relatively few (13), slender vertebrae rather than dozens of short ones (see Rieppel et al. 2010 for discussion of cervical vertebra counts in this animal). This is about half as many as some other protorosaurs had (Reippel et al. 2008), and a far cry from the vertebral counts of some dinosaurs (including birds). A low vertebral count reduces the number of heavy joints and muscle attachments in any part of the axial column, so this is a good basis to having a lightweight neck. More weight was lost through hollowing the bony core of each vertebra, a condition
Tanystropheus took so far as to need bony struts supporting the interior cavities of each vertebra. Note that there is no evidence that these bones were pneumatised, seemingly lacking openings through which airsacs could penetrate the bone walls. However, simply removing bone - one of the densest, heaviest materials in our bodies - would still throw out a lot of weight. The neck was likely lightly muscled, the mid-series vertebrae being long tubes with highly reduced processes for muscle anchorage (below) - in many respects, the vertebral bodies are similar to those of
azhdarchid pterosaurs. The role of these tubular, slender mid-series neck vertebrae is confusing at first, but they make a bit more sense once we realise that most terrestrial animals control their necks via musculature anchoring to the top and base of the neck. This was likely true for
Tanystropheus and azhdarchids because anterior and posteriormost neck vertebrae are the most complex parts of the neck skeleton, presumably reflecting attachment of more muscles in these regions. We might therefore assume their necks worked in a broadly similar to those of modern animals, weird as they are.
![]() |
| Three dimensionally preserved mid-series Tanystropheus vertebra described by Dalla Vecchia (2005). |
The seemingly lessened set of neck muscles on the
Tanystropheus neck would likely limit neck performance (i.e. the size of prey that could be lifted into the air) but, again, would facilitate weight reduction. Strong, restricting joints between the majority of the neck bones and bundles of elongate cervical ribs aided reduction of musculature further, passively resisting inter-vertebral movements which otherwise require muscle action or thick ligaments to control. Elongation of cervical ribs provides another bonus for mass reduction, this trait being linked to shifting muscles down the neck in sauropods and thus lightening the neck anterior (Taylor and Wedel 2013). With passive support structures in place, muscles operating around the neck base may have been able to support and move the neck quite easily. Indeed, areas where neck elevator muscles (such as levator scapulae and the trapezius) anchor on the shoulder blade are unusually broad and well developed in
Tanystropheus compared to other protorosaurs, and certainly a lot larger than those of long-necked aquatic animals (Araújo and Correia 2015). These are useful muscles to emphasise if you're looking to economise neck mass, being able to both lift and turn the neck by simply varying the symmetry of their activation. We also see a good set of short, robust cervical ribs and broad coracoids at the base of the neck, anchoring muscles related to strong downward neck motion (unless
Tanystropheus differed from all other tetrapods). As
Mark Robinson preemptively commented on my last post, this is starting to sound a lot like works like a mechanical crane: a lightweight, strong beam operated by long muscles and ligaments (cables and pulleys in our analogy) from a powerful, mobile base. Quite how much motion was possible at the neck base is debated, but the fact that a number of
articulated Tanystropheus specimens are preserved with distinctly elevated neck bases suggests it was more flexible than the rest of the neck, and perhaps capable of a large range of motion (Renesto 2005). This, of course, has implications for balance: if the neck could be drawn up as in fossil specimens the centre of mass would be quite far back in the body (see
the last post for more on
Tanystropheus mass distribution).
To me, this is all sounding quite sauropod- and azhdarchid-like: an economically constructed neck capable of somewhat limited, but sufficient motion to procure food in terrestrial habits, albeit food that doesn't put up too much of a fight. By contrast, the
Tanystropheus neck compares quite poorly to those of long necked aquatic animals. For one, we expect a large number of short vertebrae in long-necked aquatic animals, this permitting greater numbers of muscles working on the neck skeleton. Aquatic animal neck bones are frequently expanded to enlarge the size of muscles attaching to them, these being required to move long appendages through viscous aquatic media. This makes for a heavy neck, but perennial support provided by water renders this a moot issue. Indeed, weight is often a commodity in water rather than a problem, it providing ballast against air-filled lungs or positively buoyant tissues - it's widely known that swimming tetrapods often have entirely solid bones to increase their mass further. The neck of
Tanystropheus doesn't really match any of these features. While the number of neck bones is somewhat increased compared to other protorosaurs, the aquatic
Dinocephalosaurus has almost twice as many more - 25 - in a neck of similar proportions.
Tanystropheus neck length is mainly achieved by stretching each vertebra tremendously, the addition of another three vertebrae perhaps merely being a supportive measure to boost neck length overall (birds and sauropods do the same thing - adding more neck vertebrae is not strictly an aquatic adaptation). Reduction of neck mass in
Tanystropheus neck (and limb) bones is also at odds with expectations for an aquatic animal, the hollow cores, stiffened joints and posterior displacement of musculature being unnecessary and even disadvantageous in an aquatic setting. It's actually hard to imagine the neck of
Tanystropheus being pulled through water efficiently at all, the reduced muscle profile and long vertebrae being quite problematic and under-powered for this task. It certainly does not seem well suited to chasing and grabbing fast moving aquatic prey such as squid and fish. To me,
Tanystropheus neck anatomy just seems to make a lot more sense out of water and, given how much emphasis
Tanystropheus put on its neck tissues, I think this is a pivotal consideration when attempting to understanding its lifestyle.
Summary time: a twist in the tale?
Let's sum up these three lines of discussion. The fossil record of
Tanystropheus suggests we could find it in a variety of aquatic settings - we might average these out to say it was a denizen of coastal and nearshore environments. It clearly had a taste for seafood, although we need to be careful not to over-state what this might mean about its lifestyle. Anatomically, it seems its propulsor apparatus is best suited to non-aquatic settings and that strange neck finds overwhelmingly superior comparison to terrestrial tetrapods than it does aquatic ones. I therefore have to agree with pretty much everything said about
Tanystreopheus anatomy reflecting a 'coastal fishing' habit rather than a strictly aquatic one (e.g. Renesto 2005). I actually struggle to understand how this animal would function as a swimming predator given that its anatomy seems poorly suited to an aquatic lifestyle. Indeed, even proponents of this lifestyle acknowledge that
Tanystropheus must have been a sluggish, ineffectual aquatic predator, limited to ambushing prey from darkness (e.g. Nosotti 2007). This brings us to a twist to our
Tanystropheus story: acknowledging some big issues with the
Tanystropheus swimming hypothesis, Nosotti (2007) proposed that it was a newcomer to the aquatic realm, still carrying a lot of anatomical baggage from terrestrial ancestors. It doesn't look much like an aquatic animal because, in evolutionary terms, it's
Tanystropheus first day on the job and it's still learning the adaptive ropes for being a successful marine predator.
![]() |
| My preferred lifestyle interpretation for Tanystropheus: a Triassic croc-o-heron which snatched prey from shorelines and promontories around coastal waterways. Note the animals perched on rocks out to sea - I have no problem with this animal swimming per se (as noted above, there is reason to think it was somewhat aquatically adapted), I just don't think it lived in water. |
Personally, I find this sort of argumentation weak. It implies
Tanystropheus is somehow exempt from relationships between morphology and function well established in other animals, and seems like an excuse to dismiss contrary evidence more than it does a robust hypothesis. Above all else, this proposal suffers from elevating the proposal of aquatic
Tanystropheus to a foregone truth of its palaeobiology, and structuring other lines of evidence around that - I do not think this is not a positive approach to these sort of palaeontological investigations. I would argue contrarily that, when viewed in context of other tetrapods, the weight of evidence is
against an aquatic lifestyle, but quite consistent with a more terrestrially-based habit, and that this forms a better starting point for considering its lifestyle. To my mind,
Tanystropheus taphonomy, gut content and functional anatomy are fully consistent with it being a Triassic variant on a heron, an animal which struck at swimming prey while supported on land or in bodies of shallow water. Its smattering of minor aquatic adaptations might have been useful to cross small bodies of water, support itself on wet, soft substrates and access better fishing sites. However, the morphological onus seems to be on movement unsupported by deep water, and it might be assumed these formed a minority of adaptive pressures on
Tanystropheus anatomy. Although it is difficult to think of a perfect modern analogue for this, we might find comparable functionality and behaviours in a variety of birds, crocodylians and lizards.
OK, time to call it a day with
Tanystropheus for now, although we're not done with weird Triassic taxa yet. I've definitely caught their bug, and I'm sure we'll be spending time with several more of these fascinating oddballs in the near future. Before then, the last post I have planned this year returns us to familiar dinosaur territory, featuring an especially obscure species none of you will be familiar with. I can barely remember what it's called...
Threecerasaurus?
Trihornedabottoms? Dang - I'm sure I'll remember by next time.
This overly-long article and its artwork are made possible by Patreon
Regular readers will know that this blog and artwork is sponsored by patrons who pledge support at my
Patreon page. For as little as $1 a month you can help keep this blog going and, as a reward, you get to see a bunch of exclusive content such as prints, a discount at my art store, and bonus posts not seen anywhere else. Articles posted here also typically get some 'bonus content'. For this post, I'll be discussing the scientific and palaeoartistic reasoning behind the two
Tanystropheus paintings seen accompanying my two articles on this animal. As always, I'm very grateful to everyone who signs up!
References
- Araújo, R., & Correia, F. (2015). Plesiosaur pectoral myology. Palaeontologia Electronica, 18(1), 1-32.
- Beja, P.R. (1991). Diet of otters (Lutra lutra) in closely associated freshwater, brackish and marine habitats in south-west Portugal. Journal of Zoology (London), 225: pp. 141-152
- Dalla Vecchia, F. M. (2005). Resti di Tanystropheus, saurotterigie e “rauisuchi”(Reptilia) nel Triassico Medio della Val Aupa (Moggio Udinese, Udine). Gortania, 27, 25-48.
- Denny, M. W., & Gaines, S. D. (2007). Encyclopedia of tidepools and rocky shores (No. 1). Univ of California Press.
- Hall, C. S., & Kress, S. W. (2008). Diet of nestling Black-crowned Night-herons in a mixed species colony: implications for tern conservation. The Wilson Journal of Ornithology, 120(3), 637-640.
- Hartwick, B. (1983). Octopus dofleini. In Cephalopod Life Cycles, Vol. I: Species Accounts, ed. P.R. Boyle, pp. 277-293. Academic Press, London
- Holliday, C. M., Ridgely, R. C., Sedlmayr, J. C., & Witmer, L. M. (2010). Cartilaginous epiphyses in extant archosaurs and their implications for reconstructing limb function in dinosaurs. PLoS One, 5(9), e13120.
- Kuhn-Schnyder, E. (1959). Hand und Fuss von Tanystropheus longobardicus (Bassani). Paläontologisches Institut der Universität Zürich. 921-941.
- Li, C. (2007). A juvenile Tanystropheus sp.(Protorosauria, Tanystropheidae) from the Middle Triassic of Guizhou, China. Vertebrata PalAsiatica, 45(1), 41.
- Molnar, J. L., Pierce, S. E., & Hutchinson, J. R. (2014). An experimental and morphometric test of the relationship between vertebral morphology and joint stiffness in Nile crocodiles (Crocodylus niloticus). The Journal of experimental biology, 217(5), 758-768.
- Nosotti, S. (2007). Tanystropheus longobardicus (Reptilia, Protorosauria): Re-interpretations of the Anatomy Based on New Specimens from the Middle Triassic of Besano (Lombardy, Northern Italy). Società Italiana di Scienze Naturali e Museo Civico di Storia Naturale.
- Renesto, S. (2005). A new specimen of Tanystropheus (Reptilia, Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus. Rivista Italiana di Paleontologia e Stratigrafia, 111(3), 377-394.
- Rieppel, O., Li, C., & Fraser, N. C. (2008). The skeletal anatomy of the Triassic protorosaur Dinocephalosaurus orientalis Li, from the Middle Triassic of Guizhou Province, southern China. Journal of Vertebrate Paleontology, 28(1), 95-110.
- Rieppel, O., Jiang, D. Y., Fraser, N. C., Hao, W. C., Motani, R., Sun, Y. L., & Sun, Z. Y. (2010). Tanystropheus cf. T. longobardicus from the early Late Triassic of Guizhou Province, southwestern China. Journal of Vertebrate Paleontology, 30(4), 1082-1089.
- Taylor, M. P., & Wedel, M. J. (2013). Why sauropods had long necks; and why giraffes have short necks. PeerJ, 1, e36.
- Tschanz, K. A. R. L. (1988). Allometry and heterochrony in the growth of the neck of Triassic prolacertiform reptiles. Palaeontology, 31(4), 997-1011.
- Wild, R., (1973). Tanystropheus longobardicus Bassani: neue Ergebnisse. In: Kuhn-Schnyder, E., & Peyer, B. (eds). Die Triasfauna der Tessiner Kalkalpen, XXIII. Schweizerische Paläontologische Gesellschaft 95, 1-162.
- Witton, M. P., & Naish, D. (2008). A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS One, 3(5), e2271.